S. Nemolato,1T. Cabras,2M.U. Fanari,1 F. Cau,1M. Fraschini,3B. Manconi,2 I. Messana,2M. Castagnola,3G. Faa1
1Dipartimento di Citomorfologia, Sez. di Anatomia Patologica, Università di Cagliari; 2Dipartimento di Scienze Applicate ai Biosistemi, Università di Cagliari, Cagliari; 3Dipartimento di Scienze Cardiovascolari e Neurologiche, Università di Cagliari, Cagliari; 4Istituto di Biochimica e di Biochimica Clinica, Università Cattolica and/or Istituto per la Chimica del Riconoscimento Molecolare, CNR, Istituto Scientifico Internazionale (ISI) Paolo VI, Roma, Italy
Correspondence: Sonia Nemolato, Dipartimento di Citomorfologia, Sezione di Anatomia Pato-
logica, Università di Cagliari, Via Ospedale 46, 09124 Cagliari, Italy. Tel.: +39.070.609.2370
Fax: +39.070.657.882. E-mail: sonianemolato @libero.it - sonia.nemolato@tiscali.it
Key words: mast cells, thymosin β4 , tryptase, chymase.
Ackowledgements: we thank Mr. Ignazio Ferru for the secretarial assistance and Mrs Daniela Piga for technical support. This study was supported by “Fondazione Banco di Sardegna”, Cagliari, Sardinia, Italy.
Received for publication: 7 October 2009.
Accepted for publication: 28 November 2009.
©Copyright S. Nemolato et al., 2010
Licensee PAGEPress, Italy
European Journal of Histochemistry 2010; 54:e3
doi:10.4081/ejh.2010.e3
AbstractMast cells (MCs) are metachromatic cells that originate from multipotential hemopoietic stem cells in the bone marrow. Two distinct populations of MCs have been characterized: mucosal MCs are tryptase-positive while mast cells in skin contain tryptase and chymase. We now show that a sub-population of MCs is highly immunoreactive for thymosin β4, as revealed by immunohistochemical analyses of normal skin, normal colon mucosa and salivary gland tumors. Four consecutive serial sections from each case were immunostained for thymosin β4 (Tβ4), chymase, tryptase and stained for toluidine blue. In skin biopsies, MCs showed a comparable immunoreactivity for Tβ4, chymase and tryptase. In normal colon mucosa the vast majority of mucosal MCs expressed a strong cytoplasmic immunoreactivity for tryptase and for Tβ4, in the absence of chymase reactivity. A robust expression of Tβ4 was detected in tumor-infiltrating and peritumoral mast cells in salivary gland tumors and breast ductal infiltrating carcinomas. Tumor-infiltrating MCs also showed a strong immunoreactivity for chymase and tryptase. In this paper, we first demonstrate that normal dermal and mucosal mast cells exhibit strong expression of thymosin β4, which could be considered a new marker for the identification of mast cells in skin biopsies as well as in human tumors. The possible relationship between the degree of Tβ4 expression in tumor-infiltrating mast cells and tumor behaviour warrants further consideration in future investigations.
|
Mast cells (MCs) are metachromatic cells that originate from multipotential hemopoietic stem cells in the bone marrow, and reside in vascularized tissues where they play an important role in the innate immune system.1 The developmental pathways of mast cells within different tissues appear to be different.2,3 The observation of two distinct populations of mast cells, i.e. mucosal mast cells characterized by reactivity for chondroitin sulfate E proteoglycan4 and serosal or connective tissue-type mast cells, characterized by the ability to sinthesize heparin5 led to the concept of mast cell heterogeneity. Heterogeneity of mast cells has been reported in different tissues both in animals6 and in men.7 It has been demonstrated that mucosal mast cells differ from skin and peritoneal mast cells, even in the expression of two classes of proteases, one with trypsin-like activity, called tryptase,8 and one with a chimotrypsin-like activity, called chymase,9 which are among the major components of mast cell granules. In humans, these proteinases are differentially expressed in mast cell subtypes: mucosal MC are tryptase-positive, while mast cells in skin contain tryptase and chymase.10 These MCs subtypes are designated MCt and MCtc, respectively. The specialization of mast cells in different tissue sites has been hypothesized to be related to their involvement in physiology and pathology, which is yet poorly understood.11 MCs can also be stained with antibodies against c-kit.12 Histochemically, mast cells may be easily revealed by toluidine blue13 and by alcian blue-safranin14 stains. The latter method is able to detect a higher number of mast cells than toluidine blue staining.15 The fixation mode of tissue samples also influences the number of detected mast cells. It is commonly accepted that Carnoy’s solution has the best capacity to fix mast cell granules;16 formalin is also good and allows appropriate histochemistry and immunohistochemistry for MCs. The aim of this study is to propose the introduction of thymosin β4 (Tβ4) as a new marker for serosal, mucosal and tumor infiltrating mast cells.
In order to test Tβ4 immunoreactivity in the different subtypes of human MCs, we examined 10 surgical specimens of normal skin containing tryptase and chymase-positive serosal MCs and 10 normal colon biopsies containing tryptase-positive mucosal mast cells. Tβ4 expression in tumor-infiltrating MCs was evaluated in 4 acinic cell carcinomas of salivary glands and in 10 ductal infiltrating breast carcinomas. All 34 specimens were selected for this study by the preliminary demonstration through toluidine-blue stain of metachromatic granules-containing MCs. Positive controls included tonsil and reactive lymph nodes for chymase and tryptase and for Tβ4, respectively. All samples were fixed in 10% formalin, paraffin-embedded and routinely processed. The immunohistochemical technique was performed in a DAKO autostainer. Paraffin sections from each case were immunostained with the policlonal antibodies against thymosin β4 (Bachem-Peninsula Lab, San Carlos, CA, USA; diluted 1:500), chymase (Novocastra, Benton Lane, Newcastle, UK clone CC1, diluted 1:200) and tryptase (Novocastra, Benton Lane, Newcastle, UK clone 10D11, diluted 1:300). A LSAB2 system-HRP kit (DAKO) was used and the signal was visualized by the AEC (3-amino-9-ethylcarbazole) substrate. All samples were also stained with the toluidine-blue method.
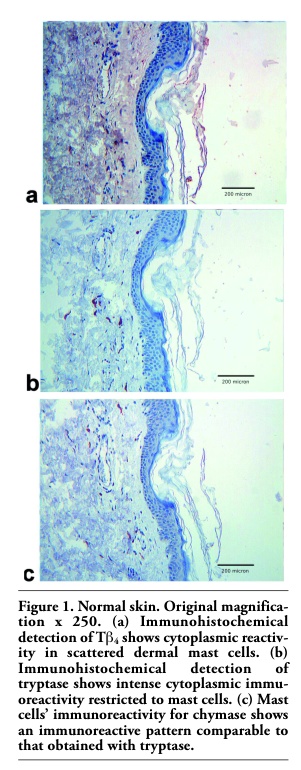
In normal skin, dermal-serosal mast cells showed substantially the same immunoreactivity for chymase (Figure 1c) and tryptase (Figure 1b). Moreover, the vast majority of MCs showed reactivity for Tβ4 (Figure 1a). Toluidin blue stain confirmed the same reactivity pattern (data not shown).
In normal colon mucosa, a different immunoreactivity pattern was detected. The vast majority of mucosal MCs expressed a strong cytoplasmic immunoreactivity for Tβ4 (Figure 2a), which paralleled the immunoreactivity for tryptase (Figure 2b). Mucosal colonic MCs did not show any significant reactivity for chymase (Figure 2c).
In salivary gland tumors, peri- and intratumoral MCs showed a comparable immunoreactivity for tryptase (Figure 3b) and chymase (Figure 3c). Immunoreactivity for Tβ4 was found only in a subset of intratumoral mast cells. (Figure 3a).
In breast ductal infiltrating carcinomas, tumor infiltrating and peritumoral MCs showed a strong reactivity for Tβ4 and tryptase; immunoreactivity for chymase was detected only in a subset of MCs. The same pattern of reactivity was found in MCs surrounding fibrocystic changes (Figure 4 a, b, c).
|
Figure 1. Normal skin. Original magnification x 250. (a) Immunohistochemical detection of Tb4 shows cytoplasmic reactivity in scattered dermal mast cells. (b) Immunohistochemical detection of tryptase shows intense cytoplasmic immu-oreactivity restricted to mast cells. (c) Mast cells’ immunoreactivity for chymase shows an immunoreactive pattern comparable to that obtained with tryptase. |
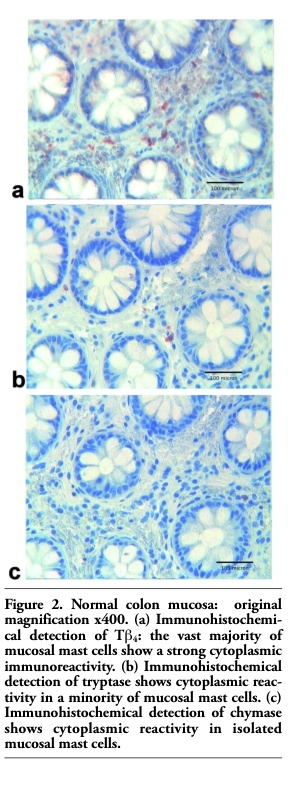 |
Figure 2. Normal colon mucosa: original magnification x400. (a) Immunohistochemi-cal detection of Tb4: the vast majority of mucosal mast cells show a strong cytoplasmic immunoreactivity. (b) Immunohistochemical detection of tryptase shows cytoplasmic reactivity in a minority of mucosal mast cells. (c) Immunohistochemical detection of chymase shows cytoplasmic reactivity in isolated mucosal mast cells. |
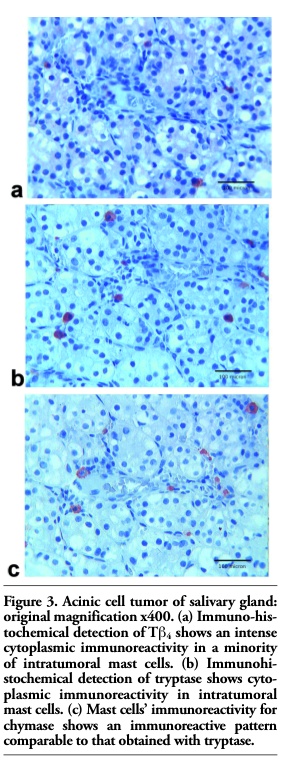 |
Figure 3. Acinic cell tumor of salivary gland: original magnification x400. (a) Immuno-histochemical detection of Tb4 shows an intense cytoplasmic immunoreactivity in a minority of intratumoral mast cells. (b) Immunohi- stochemical detection of tryptase shows cytoplasmic immunoreactivity in intratumoral mast cells. (c) Mast cells’ immunoreactivity for chymase shows an immunoreactive pattern comparable to that obtained with tryptase. |
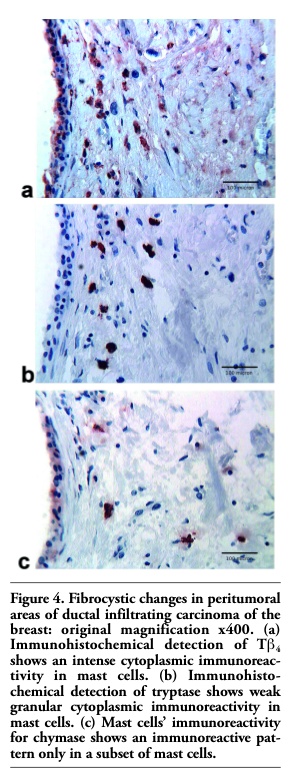 |
Figure 4. Fibrocystic changes in peritumoral areas of ductal infiltrating carcinoma of the breast: original magnification x400. (a) Immunohistochemical detection of Tb4 shows an intense cytoplasmic immunoreactivity in mast cells. (b) Immunohisto- chemical detection of tryptase shows weak granular cytoplasmic immunoreactivity in mast cells. (c) Mast cells’ immunoreactivity for chymase shows an immunoreactive pattern only in a subset of mast cells. |
In this paper, we first demonstrate that normal dermal (serosal) and mucosal mast cells exhibit strong expression of thymosin β4. Tβ4 expression was also found in tumor-infiltrating and peritumoral mast cells in salivary gland and in breast tumors. These observations extend the knowledge regarding the immunophenotypic profile of mast cells and provide a useful new marker for the identification of scattered mast cells in skin, colon mucosa as well as in tumors. The co-expression or separate expression of chymase and tryptase is currently used as a specific immunophenotypic profile to distinguish and characterize, on tissue sections, different subtypes of mast cells.10 According to our results, Tβ4 could be considered as a marker for MCs in this context, showing a degree of immunoreactivity comparable in sensitivity to chymase and tryptase. In this study Tβ4 demonstrated to stain both serosal and mucosal MCs, paralleling the sensitivity of tryptase in revealing tissue MCs.
In addition, in this study we found a pattern of reactivity for Tβ4 which could be useful to better characterize, on tissue sections, a new subtype of tumor infiltrating and peritumoral MCs. In salivary gland tumors, Tβ4 was detected only in a part of tumoral MCs, while reactivity for chymase and tryptase was found in all MCs. A different protein reactivity pattern for Tβ4 was found in intra- and peri-tumoral MCs in infiltrating ductal carcinoma of the breast. In this contest, MCs showed the highest level of reactivity for Tβ4 and tryptase, whereas only a minority of MCs were positive for chymase. All together, these findings show a previously unreported heterogeneity of tumor infiltrating MCs, and introduce Tβ4 as a useful marker, in association with chymase and tryptase for the identification of new subtypes of MCs.
As far as the role of mast cells inside and around malignant neoplasms is concerned, the demonstration of strong Tβ4 expression in intratumoral MCs can contribute to explain their function as effectors of the innate immune systems in the surveillance of tumor progression. A number of experimental, biochemical and microarray studies have already documented the presence of a high level of expression of Tβ4 in experimental and human tumors.17,18 In particular, the role of Tβ4 has been analyzed in relation to tumor metastasis,19,20,21 in human lung cancer,22 and in human colorectal cancer.23 Recently, an immunohistochemical study on Tβ4 expression in human breast cancer revealed a low reactivity for the peptide in cancer cells, while intense reactivity for Tβ4 was detected in other cells of the tumor microenvironment.24 On the basis of our data, we may hypothesize that Tβ4- positive peritumoral cells observed in that study could be identified as MCs.
These data taken together, one may pose the question whether the levels of Tβ4 recorded in extracts of tumor samples in previous studies reflected levels in tumor cells or in the tumour microenvironment. Our data give an answer to this question, and clearly indicate mast cells as partly responsible for the connection between Tβ4 and tumour progression. The finding of a strong reactivity for Tβ4 in tumour-infiltrating mast cells lays stress on the hypothesis that Tβ4 may constitute a local paracrine mediator which could be relevant to our understanding of cellular cross-talk within the tumor micro-environment.24 Tβ4 has been shown to exert anti-inflammatory and cytoprotective functions, by suppressing secretion of the pro-inflammatory cytokine, IL8 and by protecting cells against apoptosis induced by tumor necrosis factor-a.25 Moreover, in a murine corneal model, Tβ4 potently inhibited neutrophil infiltration and significantly decreased the expression of pro-inflammatory cyto-kines.26 These data taken together, we may speculate that, because of its multifunctional roles in protecting cells against apoptosis25 and in stimulating neoangiogenesis,27,28,29 Tβ4 released by mast cells in the tumor microenvironment could significantly contribute to cancer cell survival. As a consequence, Tβ4-producing MCs could represent a new molecular target for antitumoral strategies30 in different human tumors.
Finally, the expression of Tβ4 in the cytoplasm of tumor-infiltrating mast cells reported in this study extends our knowledge regarding the immunophenotypic profile of mast cells and can significantly contribute to our understanding of cross-talk between cells of the immune system and tumour cells. The possible relationship between the degree of mast cell infiltration and tumor behaviour warrants further consideration in future investigations.
1. Rao KN, Brown MA. Mast cells. Multi-faceted immune cells with diverse roles in health and disease. Ann N.Y. Acad Sci 2008;1143:83-104. [PubMed]
2. Kitamura Y, Kasugai T, Arizono N, Matsuda H. Development of mast cells and basophils: processes and regulation mechanisms. Am J Med Sci 1993;306:185-91. [PubMed]
3. Barret K, Pearce F. Heterogeneity of mast cells. In: Uvnas B, ed. Histamine and histamine antagonist, Springer-Verlag, Berlin 991, pp. 93-116.
4. Razin E, Ihle JN, Seldin D et al. Interleukin 3: a differentiation and growth factor for the mouse mast cell that contains condroitin sulfate E proteoglycan. J Immunol 1984;132:1479-86. [PubMed]
5. Tsai M, Takeishi T, Thompson H et al. Induction of mast cell proliferation, maturation, and heparin synthesis by the rat c-kit ligand, stem cell factor. Proc Natl Acad Sci 1991;88:6382-6.[PubMed]
6. Barret K, Szucs E, Metcalfe D. Mast cells heterogeneity in higher animals: a comparison of the properties of autologous lung and intestinal mast cell from non-human primates. J Immunol 1986;137: 2001-8. [PubMed]
7. Befus D, Goodacre R, Dyck N, Bienenstock J. Mast cell heterogeneity in man. I. Histologic studies of the intestine. Int Arch Allergy Appl Immunol 1985;76: 232-6. [PubMed]
8. Schwartz LB, Lewis RA, Austen KF. Tryptase from human pulmonary mast cells. Purification and characterization. J Biol Chem 1981;256:11939-43. [PubMed]
9. Schechter NM, Choi JK, Slavin DA et al. Identification of a chymotrypsin-like proteinase in human mast cells. J Immunol 1986;137:962-70. [PubMed]
10. Caughey GH. New developments in the genetics and activation of mast cell proteases. Mol Immunol 2002;38:1353-7.[PubMed]
11. Beaven MA. Our perception of the mast cell from Paul Ehrlich to now. Eur J Immunol 2009;39:11-25. [PubMed]
12. Edwards ST, Cruz AC, Donnelly S, et al. c-Kit immunophenotyping and metalloproteinase expression profiles of mast cells in interstitial lung diseases. J Pathol 2005; 206:279-90.[PubMed]
13. Pearce A. Histochemistry 2th ed. J&A Churchill Ltd, 1960.
14. Csaba G. Alcian blue-safranin method for mast cells. In: JD Bancroft and Steven A eds. Theory and practice of histological techniques. Livingstone, New York, 1990, p 639.
15. Vodenicharov A, Leiser R, Gulubova M, Vlaykova T. Morphological and immunocytochemical investigations on mast cells in porcine ureter. Anat Histol Embryol 2005;34:343-9. [PubMed]
16. Frangogiannis N, Burns A, Michael L, Entman M. Histochemical and morphological characteristics of canine cardiac mast cells. Histochem J 1999; 31:221-9.[PubMed]
17. Chen C, Li M, Yang H et al. Role of thymosins in cancer and other organ systems. World J Surg 2005;29:264-70. [FullText]
18. Huff T, Muller CSG, Otto AM et al. Beta-thymosins, small acidic peptides with multiple functions. Int J Biochem Cell Biol 2001;33:205-20. [PubMed]
19. Cha HJ, Jeong KJ, Kleinman HK . Role of thymosin beta4 in tumor metastasis and angiogenesis. J Natl Cancer Inst 2003;95: 1674-80. [PubMed]
20. Kobayashi T, Okada F, Fujii N et al. Thymosin beta4 regulates motility and metastasis of malignant mouse fibrosarcoma cells. Am J Pathol 2002;160:869-82. [PubMed]
21. Wang WS, Chen PM, Hsiao HL, Ju SY. Overexpression of the thymosin beta4 gene is associated with malignant progression of SW480 cancer cells. Oncogene 2003;22:3297-306. [PubMed]
22. Ji P, Diederichs S, Wang W et al. MALAT-1 a novel noncoding RNA and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003;22:6087-97. [PubMed]
23. Wang WS, Chen PM, Hsiao HL et al. Over-expression of the thymosin beta4 gene is associated with increased invasion of SW480 colon carcinoma cells and the distant metastasis of human colorectal carcinoma. Oncogene 2004;23:6666-71. [FullText]
24. Larsson LI, Holck S. Occurrence of thymosin beta4 in human breast cancer cells and in other cell types of the tumor microenvironment. Hum Pathol 2007;38: 114-9.[PubMed]
25. Reti R, Kwon E, Qiu P et al. Thymosin beta4 is cytoprotective in human gingival fibroblasts. Eur J Oral Sci 2008;116:424-30.[PubMed]
26. Sosne G, Chan CC, Thai K et al. Thymosin beta 4 promotes corneal wound healing and modulates inflammatory mediators in vivo. Exp Eye Res 2002;74:293-9.[PubMed]
27. Malinda KM, Goldstein AL, Kleinman HK. Thymosin beta 4 stimulates directional migration of human umbilical vein endothelial cells. FASEB J. 1997;11:474-81.[PubMed]
28. Grant DS, Rose W, Yaen C et al. Thymosin beta4 enhances endothelial cell differentiation and angiogenesis. Angiogenesis 1999;3:125-35.[PubMed]
29. Smart N, Rossdeutsch A, Rile PR. Thymosin Beta4 and angiogenesis: modes of action and therapeutic potential. Angiogenesis 2007;10:229-41. [PubMed]
30. Goldstein AL. Thymosin beta4: a new molecular target for antitumor strategies. J Natl Cancer Inst 2003;95:1646-7.[PubMed]
[TOP]