 |
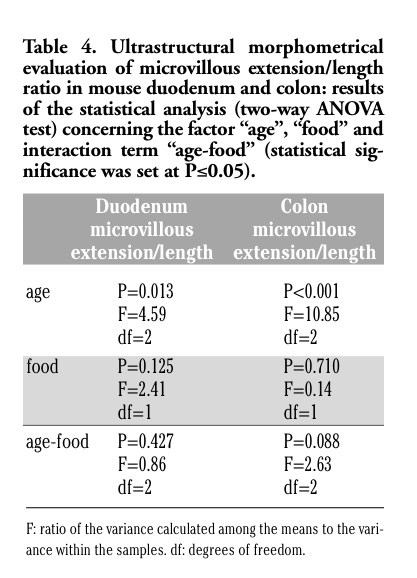
Table 1. Morphometrical analyses in duodenum and colon of control and GM-fed mice (means±SE). |
S. Battistelli,1 B. Citterio,2 B. Baldelli,1C. Parlani,2 M. Malatesta3
1Department of Biomolecular Sciences, Clinical Biochemistry Section, University of Urbino, Italy; 2Department of Biomolecular Sciences, Section of Toxicological, Hygienistic and Environmental Sciences, University of Urbino, Italy; 3Department of Neurological, Neuropsychological, Morphological and Motor Sciences, Anatomy and Histology Section, University of Verona, Italy
Correspondence:Dr. Serafina Battistelli, Department of Biomolecular Sciences, Clinical Biochemistry Section, University of Urbino, Via Ubaldini 7 - 61029 Urbino (PU), Italy.
Tel. +39.0722.351084 - Fax +39.0722.322370.
E-mail: serafina.battistelli@uniurb.it
Key words: diet, genetically modified soybean, intestine, goblet cell mucins, coliform bacteria.
Acknowledgments: we are particularly grateful to Dr. F. Marcheggiani for his skilful technical assistance and to Dr. F. Marinelli for his valuable help for statistical analysis. This study was supported by the Agenzia Servizi Settore Agroalimentare delle Marche, Italy; this sponsor did not have any involvement in the study processing nor in the decision to submit the paper for publication.
Received for publication: 27 April 2010.
Accepted for publication: 24 June 2010.
This work is licensed under a Creative Commons Attribution 3.0 License (by-nc 3.0).
©Copyright S. Battistelli et al., 2010
Licensee PAGEPress, Italy
European Journal of Histochemistry 2010; 54:e36
doi:10.4081/ejh.2010.e36
AbstractDiet can influence the structural characteristics of both small and large intestine. In this study, we investigated the duodenum and colon of mice fed on genetically modified (GM) soybean during their whole life span (1-24 months) by focusing our attention on the histological and ultrastructural characteristics of the epithelium, the histochemical pattern of goblet cell mucins, and the growth profile of the coliform population. Our results demonstrate that controls and GM-soybean fed mice are similarly affected by ageing. Moreover, the GM soybean-containing diet does not induce structural alterations in duodenal and colonic epithelium or in coliform population, even after a long term intake. On the other hand, the histochemical approach revealed significant diet-related changes in mucin amounts in the duodenum. In particular, the percentage of villous area occupied by acidic and sulpho-mucin granules decreased from controls to GM-fed animals, whereas neutral mucins did not change.
|
Diet can influence the characteristics of the gastrointestinal tract since the intestinal mucous membrane is directly in contact with food and absorbs the substances produced by digestion. In particular, it has been reported that the diet may affect both small and large intestine in terms of mucosal architecture, villous height and crypt depth, epithelial cell proliferation and other features.1,2 Moreover, it is known that diet and the histochemical characteristics of goblet cell mucins and/or mucous membrane are strictly correlated.3
While extensive literature exists on the changes induced on gut histology by different dietary factors such as fibres, pectin, cellulose, nucleotides, etc.,4 scarce data have so far been reported on the effects of diets containing genetically modified (GM) plants on the small and large intestine, especially with regard to the possible long term consequences.5,6 In particular, the high sensitivity of goblet cell mucins to different physiological and pathological conditions3 makes them suitable markers for monitoring diet effects on the intestine.
To this aim, in this study we investigated some features of duodenum and colon of mice fed on GM soybean during their whole life span (1-24 months) by focusing our attention on the histological and ultrastructural characteristics of the epithelium, the histochemical pattern of goblet cell mucins and the growth profile of the coliform population.
Twenty-four female Swiss mice were used. Twelve of them were fed ad libitum on a standard laboratory chow containing 14% GM soybean obtained by the insertion of the bacterial CP4 EPSPS gene conferring a high level of tolerance to glyphosate, the active ingredient of the herbicide Roundup (GTS 40-3-2).7 In parallel, twelve mice (controls) were fed on a diet containing the same nutrition components with the exclusion of the GM soybean, which was replaced by a commercial, non-GM soybean.
The mice were weighed before being killed by cervical dislocation when they were 1, 12 and 24 months old. Samples of the duodenum and colon were quickly removed and immediately processed for light and electron microscopy.
For histology, morphometry and histochemistry at light microscopy, samples were fixed with 4% neutral buffered formalin and embedded in paraffin wax. Five µm-thick cross sections of duodenum and colon were either stained with hematoxylin-eosin for morphological examinations or processed for carbohydrate histochemistry using either the periodic acid - Schiff (PAS) reaction for neutral mucins, or the Alcian blue at pH 2.5 for acidic mucins, or the high iron diamine-alcian blue technique for sulpho-mucins according to the routine methods. For each method, ten sections from both duodenum and colon were stained. Observations were performed by an Olympus BX51 light microscope. Morphometrical analyses were performed on micrographs (20X) by using the image analysis systems CELL-B for Windows XP. For each sample, 30 intact and well-oriented villi and/or crypts were evaluated. The external (serosal surface) and internal (luminal face) perimeters of the intestinal wall were measured and their ratio was calculated in order to evaluate the extent of villi and/or crypts independently of the bowel size. The height and total area of villi and/or crypts were also measured (the mean area±SE measured per animal was 1062379.2±26939.1 µm2 for duodenal villi, 143639.8±1904.5 µm2 for duodenal crypts and 359282.3±3542.4 µm2 for colonic crypts); in addition, the villous and/or crypt area occupied by granules stained for neutral, acidic or sulpho-mucins were measured, and their per cent area was calculated with respect to the total area.
For ultrastructural morphological and morphometrical evaluations, duodenum and colon samples were fixed in a mixture of 2.5% glutaraldehyde and 2% paraformaldehyde in 0.1M Sörensen phosphate buffer, pH 7.4 for 3 h, post-fixed with 1% OsO4 at 4°C for 1 h and embedded in Epon. Ultrathin sections were stained with lead citrate and observed in a Philips Morgagni TEM equipped with a Megaview II camera for digital image acquisition. Ten electron micrographs (20,000X) of both duodenal and colonic enterocyte surface were taken for each animal and the microvillous extension/length ratio was evaluated by using the image analysis systems Image Pro-Plus for Windows 95.
For evaluation of coliform bacteria, faecal samples were collected weekly. They were aseptically weighed, 9 mL of PBS/g of faeces (vol/wt) were added, and the mixture was incubated for 15 min at 25°C. For the detection of total coliform count, 8-fold dilutions were carried out and 1 mL of each dilution was surface plated in triplicate on MacConkey Agar (Oxoid) and incubated at 37°C for 24 h. The following identification tests were performed: Lactose fermentation, Triple Sugar Iron Agar (TSI) (OXOID) and API 20 E (BioMérieux).
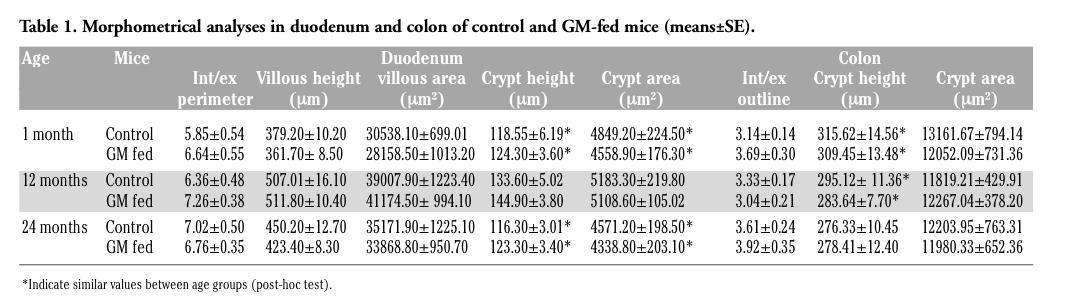
Since all parameters analysed may undergo modifications due to either the age of the animals or the diet or both of them, statistical comparisons were performed by the two-way ANOVA test to evaluate the factors “age” and “food” as well as the interaction term between these two factors. Furthermore, in order to determine which pairs of samples tended to differ for the age factor, the Tukey’s HSD test for multiple comparisons was used (post-hoc test).
At sacrifice, mouse body weight varied from 21 to 38 g, without significant differences between control and GM-soybean fed animals of the same age. Neither macroscopic alterations nor pathologic lesions were observed in the whole intestine or in other organs at the time of death. According to previous studies,5 this demonstrates that the GM soybean-containing diet does not affect the gross anatomy or the histopathological features, even after a long lasting intake.
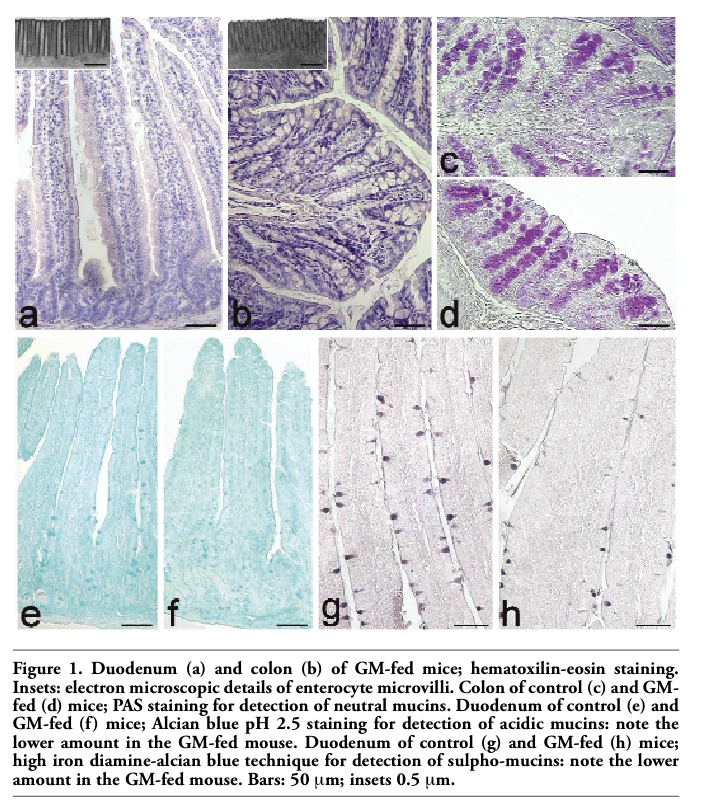
The epithelium covering the free surface of the mucous membrane was simple columnar and no structural alteration was observed in any animal; the villi showed finger-like, tongue-like or branched shapes, without apparent difference between control and GM soybean-fed mice (Figure 1 a,b). Duodenal and colonic enterocytes revealed similar ultrastructural features in control and GM-fed animals: the cells appeared markedly elongated and polarised, with a basal nucleus and an apical brush border (insert in Figure 1 a,b). No evident morphological modification was observed in nuclei or in cytoplasmic organelles, i.e., the rough and smooth endoplasmic reticulum, Golgi complex and mitochondria, between the two animal groups in both duodenum and colon (not shown). These observations demonstrate that the glyphosate-tolerant GM soybean administered to mice for 1, 12 and 24 months does not induce structural alterations in duodenal and colonic epithelium, as seen at light and electron microscopy. The morphological data were confirmed by the morphometrical results (Tables 1 and 2), which excluded any significant difference due to the food factor (Tables 3 and 4).
As for the structural changes related to age observed in both duodenum and colon of both animal groups, they are consistent with the results of several studies describing the effects of aging on the intestine:8 since the height of villi and the extension of microvilli in the small intestine are related to the digestive and adsorptive capability,9 the low values observed in 1 month-old mice (which cannot be considered as mature mice yet) could be related to the incomplete development of the digestive tract, while the reduction found in 24 month-old mice could be associated with a decreased intestinal functionality.
On the other hand, the histochemical approach revealed some age- and diet-related differences.
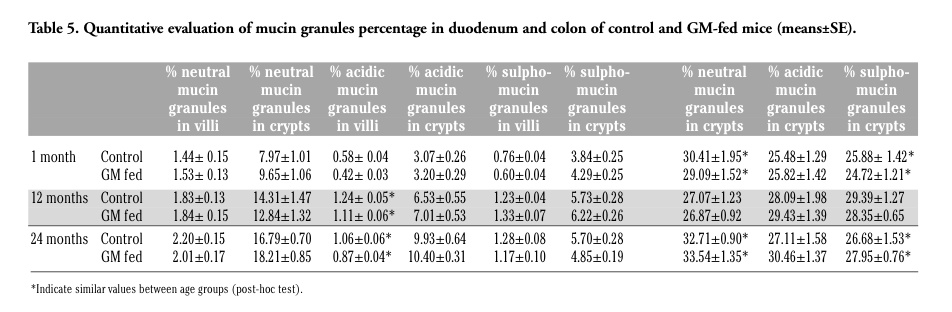
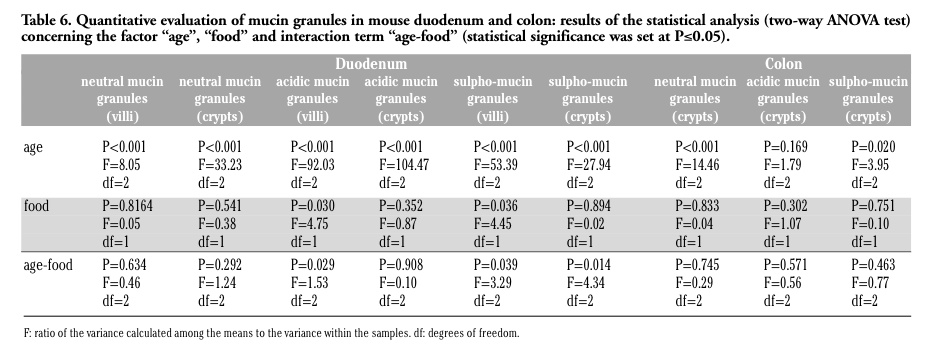
Neutral mucins increased with increasing age in both animal groups and there was a similar staining pattern, without significant differences in GM-fed and control mice (Figure 1 c,d) (Tables 5 and 6). The amount of acidic mucins increased from 1 to 12 months old control mice, while decreasing in the duodenal villi of 24 months old animals: the low amount of acidic mucins in young mice could again be related to their still incomplete development of the intestine, whereas the decrease in acidic and sulpho-mucins (which are particularly efficient in protecting against bacterial penetration),1 and the parallel increase in the less viscous neutral mucins in 24 months old mice is consistent with the reduced resistance to intestinal infections typical of the old age.8,10
The modification pattern of acidic and sulpho-mucins is similar in control and GM-fed mice, at all the ages considered. However, in the duodenal villi of GM-fed animals, a lower amount of acidic mucins (Figure 1 e,f) and sulpho-mucins (Figure 1 g,h) was observed in comparison to controls. This was confirmed by morphometrical and statistical analyses (Tables 5 and 6) and by the significant interaction term between the factors “age” and “food”.
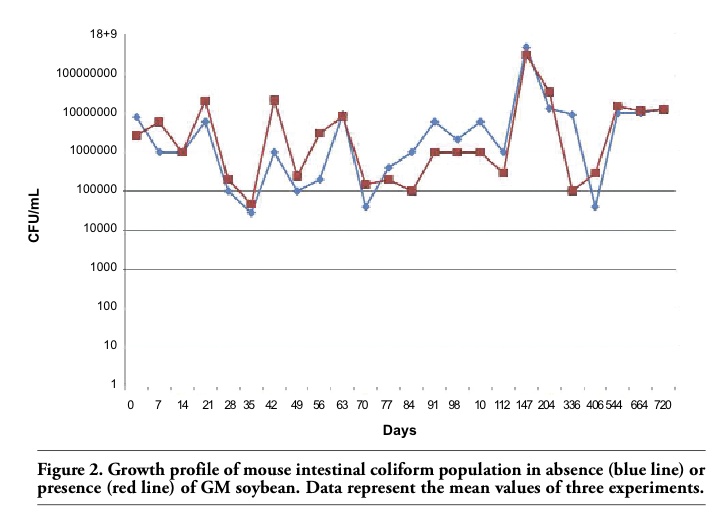
Changes in the intestinal mucins are directly linked to the dynamic equilibrium between their biosynthesis by goblet cells and their degradation within the lumen by the microflora, and the levels of mucins may vary in the epithelium according to the diet and to bacteria inhabiting the digestive tract.3 As our data on coliforms demonstrated similar growth profiles in the two animal groups (Figure 2), it is likely that the differences observed in the amount of mucins might be related to the effect of the GM-containing diet. However, at the present state of the research, the factor(s) responsible for the modifications in duodenal mucin production are still to be identified: actually, the GM soybean used in this study has been treated with the herbicide Roundup to which it had been rendered tolerant, and it is impossible to choose whether the differences observed may be related to the GM soybean itself or to herbicide residues possibly present in the feed. In fact, studies on the effects of Roundup or its active ingredient, glyphosate, have demonstrated alterations of the intestinal mucosa in different animal.5 Anyhow, acidic and sulpho-mucins are characterised by a higher viscosity compared to the neutral mucins, thus conferring a higher degree of protection to the intestinal surface,1 and their decrease could make the intestine less resistant to infections. Interestingly, only the duodenal tract appears to be affected by the diet: it is tempting to speculate that the food components cannot affect the colon epithelium because they have already been mostly degraded by the digestive enzymes when they reach this intestine tract.
|
|
|
|
|
|
|
|
1. Hedemann MS, Eskildsen M, Laerke HN, Pedersen C, Lindberg JE, Laurinen P, et al. Intestinal morphology and enzymatic activity in newly weaned pigs fed contrasting fiber concentrations and fiber properties. J Anim Sci 2006;84:1375-86. [Pubmed]
2. Hartke JL, Monaco MK, Wheeler MB, Donovan SM. Effect of a short-term fast intestinal disaccharidase activity and villus morphology of piglets suckling insulin-like growth factor-I transgenic sows. J Anim Sci 2005;83:2404-13.[Pubmed]
3. Montagne l, Pluske JR, Hampson DJ. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim Feed Sci Technol 2003;108:95-117.[Abstract]
4. Hedemann MS, Mikkelsen LL, Naughton PJ, Jensen BB. Effect of feed particle size and feed processing on morphological characteristics in the small and large intestine of pigs and on adhesion of Salmonella enterica serovar Typhimurium DT12 in the ileum in vitro. J Anim Sci 2005;83:1554-62.[Pubmed]
5. Malatesta M. Animal feeding trials for assessing GMO safety: answers and questions. CAB Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources 2009;4,068:1-13.
6. Séralini GE, Cellier D, de Vendomois JS. New analysis of a rat feeding study with a genetically modified maize reveals signs of hepatorenal toxicity. Arch Environ Contam Toxicol 2007;52:596-602.[Pubmed]
7. Padgette SR, Kolacz KH, Delannay X, Re DB, LaVallee BJ, Tinius CN, et al. Development, identification and characterization of a glyphosate-tolerant soybean line. Crop Sci 1995;35:1451-61.[Abstract]
8. Trbojević-Stanković JB, Milićević NM, Milosevićć DP, Despotovićć N, Davidovićć M, Erceg P, et al. Morphometric study of healthy jejunal and ileal mucosa in adult and aged subjects. Histol Histopathol 2010;25:153-8.[Pubmed]
9. Pluske JR, Hampson J, Williams H. Factors influencing the structure and function of the small intestine in the weaned pig. A review. Livest Prod Sci 1997;51:215-36. [Abstract]
10. Lindèn SK, Florin TH, McGuckin MA. Mucin dynamics in intestinal bacterial infection. PLoS One 2008;3:3952-65. [Pubmed]
[TOP]