S. Ćirović,1 J. Marković-Lipkovski,1 J. Todorović,2 J. Nešović-Ostojić,2 M. Jović,3 S. Ilić,3 S. Tatić,1 D. Čemerikić2
1Institute of Pathology, Medical Faculty, Belgrade, Serbia; 2Institute of Pathological Physiology, Medical Faculty, Belgrade, Serbia; 3Center for Pathology and Forensic Medicine, Military Medical Academy, Belgrade, Serbia
Correspondence: Jasmina Marković-Lipkovski
Institute of Pathology, Faculty of Medicine, University of Belgrade Dr Subotića 1/I, 11000 Belgrade, Serbia.
E-mail: acal@matf.bg.ac.rs
Key words: KCNQ1 K+ channel; rana esculenta; frog kidney; immunohistochemistry.
Acknowledgements: this work was supported by grants No. 145085 and 145034 from the Ministry of Science and Technological Development of the Republic of Serbia.
Received for publication: 7 October 2009.
Accepted for publication: 7 January 2010.
©Copyright S Ćirović et al., 2010
Licensee PAGEPress, Italy
European Journal of Histochemistry 2010; 54:e7
doi:10.4081/ejh.2010.e7
AbstractThe aim of this study was to evaluate KCNQ1 K+ channel expression in the frog kidney of Rana esculenta. KCNQ1 K+ channel, also known as KvLQT1, is the pore forming a-subunit of the IKs K+ channel, a delayed rectifier voltage-gated K+ channel, which has an important role in water and salt transport in the kidney and gastrointestinal tract. The expression of KCNQ1 K+ channel along tubular epithelium differs from species to species. In the present study the expression of KCNQ1 K+ channel in the frog kidney has been demonstrated by immunohistochemistry. The presence of KCNQ1 K+ channel was demonstrated in the epithelial cells of distal convoluted tubule and collecting duct. However, the pattern of expression of KCNQ1 K+ channel differs between distal convoluted tubules and collecting duct. All epithelial cells of distal convoluted tubules revealed basolateral expression of KCNQ1 K+ channel. On the contrary, only the single cells of collecting duct, probably intercalated cells, showed diffuse cell surface staining with antibodies against KCNQ1 K+ channel. These findings suggest that KCNQ1 K+ channel has cell-specific roles in renal potassium ion transport. |
K+ channels form a large family of membrane proteins that are expressed in epithelial cells and have important functions, such as maintenance of membrane potential, volume regulation, recycling and secretion of K+.1,2 KCNQ1 K+ channels are expressed in several epithelial tissues and the heart and regulate key physiological functions.3 KCNQ1 K+channels are made up of four α-subunits, each containing six transmembrane segments, a pore loop constituting the central part of the pore holding for potassium selectivity filter and intracellular N- and C-termini.4 A K+ channel α subunit KCNQ1, formerly known as KvLQT1,5 associates with KCNE1 (minK channel, IsK), a K+ channel β subunit, expressed in the apical cell membrane of mammalian proximal renal tubules,6,7 the heart and a number of epithelial cells,8 to produce a current that resembles the slow delayed rectifier current (IKs) in cardiac myocytes.9,10 Recently has been published that KCNQ1 together with KCNE1 are involved in early left-right polarization in frog embryos.11 An important role of the luminal KCNQ1 K+ channel in proximal tubules as the driving force for electrogenic Na+ and substrate reabsorption has been presented in the KCNQ1-deficient mice (KCNQ1-/-).12 Our previous results confirmed the absence of KCNQ1 K+ secretory fluxes in the proximal cells of frog kidney with RT-PCR analysis which revealed that KCNQ1 mRNA is not expressed in the proximal tubules of frog kidney at a detectable level, as used specimens of the frog kidney tissue for RT-PCR analysis were from lateral upper pole of the frog kidney which contained proximal tubules.13
The presence of KCNQ1 in mouse kidneys distal and collecting tubules but not in proximal tubules has been demonstrated by immunofluorescence, in situ hybridization8 and by RT-PCR and immunohistochemistry.14 However, there are inconsistent data about renal expression of KCNQ1 K+ channel in different animals, even within the same species. The purpose of this study was to determine the tubule location of KCNQ1 K+ channel protein in frog kidney applying antibody against the carboxyl terminus of human KCNQ1 protein.
Rana esculenta (<50 g) of both sexes (adult frog) were kept in aquariums at 4°C. After anaesthesia with tricaine methanesulfonate (1.5 g/L adjusted to pH 7.0), the kidneys from five frogs were carefully removed from the animals and placed in 10 mL of Ringer solution. Amphibian Ringer solution of the following composition (in mmol/L) was used: NaCl 90, KCl 2.5, NaHCO3 10, Na H2 PO4 0.5, CaCl2 1.8, MgCl2 1, glucose 2.2, dextran (mol. wt. 80 000) 15 g/L and heparin 2000 U/L. The Ringer solution was equilibrated with 1% CO2 - 99% O2 to pH 7.6. The kidneys were doubly perfused with aortic and portal vein perfusions, maintained using hydrostatic pressure (50 cm H2O for aortic perfusion, 30 cm H2O for portal vein perfusion). The doubly perfused kidneys were then cut. The kidneys were harvested and kept in 4% paraformaldehyde solution at room temperature, at which point they were embedded in paraffin, using the Leica ASP300 automated paraffin wax tissue processor. Five-micrometer sections were cut and blocked with 0.05M Tris-EDTA, pH 9.0 for 20 min in microwave oven. Then, sections were incubated with KCNQ1 (C-20) antibody (catalogue no. sc-10646; Santa Cruz Biotechnology) at 1:50 dilution dissolved in PBS (Phosphate Buffered Saline, pH 7.4, 50 mM NaH2PO4, 150 mM NaCl, 0.1% Tween 20, and 3% BSA) for 60 minutes. KCNQ1 is a goat polyclonal antibody raised against a peptide mapping at the C-terminus of KCNQ1 of human origin. We used this antibody on frog renal tissues since, according to the NCBI database (http://www. ncbi.nlm.nih.gov/pubmed/), KCNQ1 protein sequence in Homo sapiens (Gene ID 3784) and KCNQ1-A protein sequence in Xenpous laevis (Gene ID 373746) have three equal conserved domains, and they match in 86% and are identical in 77%.
Localization of KCNQ1 was accomplished using immunoperoxidase procedures as we have done previously.15 Universal Dako ChemMeteTM EnVision kit (catalogue no. K5007, Dako Corporation, CA93013 USA) and 3-amino-9-ethylcarbazole (AEC, catalogue no. K 3469, Ready-to-use, Dako Corporation, CA93013 USA), were used for detection and visualization of KCNQ1 expression. After that sections were analyzed on Nikon Coolscope microscope. Controls in immunostaining procedure were obtained by replacement of the primary antibody with PBS or by blocking antibodies (sc-10646 P) Santa Cruz.
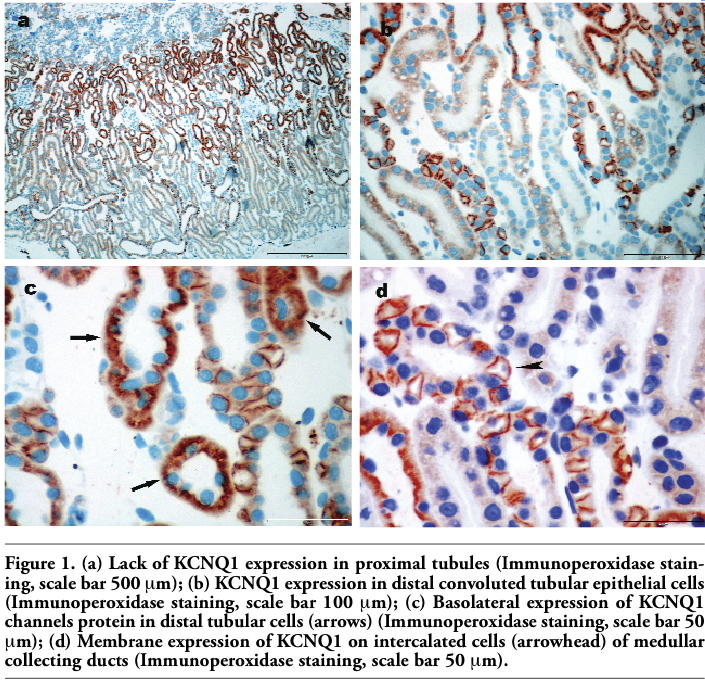
In the present study we analyzed KCNQ1 K+ channel expression in the frog kidney by means of immunohistochemistry staining. The pattern of immunoreactivity was consistent on all slides. Observation by light microscopy on low magnification (Figure 1a) revealed predominant expression of KCNQ1 K+ channel in the tubules in the medulla, on single cells in pyramids and almost absence in the cortex. On higher magnification it was clear that proximal tubular cells were negative for KCNQ1 K+ channel immunoreactivity on all analyzed renal tissues. In the medulla mostly distal convoluted tubular epithelial cells and particular cells of collecting ducts exhibited intense staining for KCNQ1 K+ channel (Figure 1b). Distal tubular cells which could be recognized by their typical morphology, flattened epithelium and basically localized nuclei, revealed intense basolateral staining that extended deep into the cells (Figure 1c). On the contrary, only single cells of collecting ducts, very likely intercalated cells considering their appearance and position within duct, showed diffuse membrane staining (Figure 1d). Labelling was not observed in sections from frog kidney when the primary antibody was omitted or blocking antibody was performed (see on-line supplementary files).
|
Figure 1. (a) Lack of KCNQ1 expression in proximal tubules (Immunoperoxidase staining, scale bar 500 μm); (b) KCNQ1 expression in distal convoluted tubular epithelial cells (Immunoperoxidase staining, scale bar 100 μm); (c) Basolateral expression of KCNQ1 channels protein in distal tubular cells (arrows) (Immunoperoxidase staining, scale bar 50 μm); (d) Membrane expression of KCNQ1 on intercalated cells (arrowhead) of medullar collecting ducts (Immunoperoxidase staining, scale bar 50 μm). |
In the present study, we clearly show the absence of KCNQ1 K+ channel expression by mean of immunomorphology, on proximal tubule epithelial cells. This finding confirmed our previous results concerning the absence of KCNQ1 K+ secretory fluxes in the proximal cells of frog kidney, where we investigated the effects on rapid depolarization and slow repolarization of the peritubular membrane potential after luminal addition of substrates for Na+ coupled transport with KCNQ1 K+ channel blockers. Also, RT-PCR analysis in the same study revealed that KCNQ1 mRNA is not expressed in the proximal tubules of frog kidney at a detectable level, although revealed a detectable KCNQ1 expression in the frog heart.13 Used specimens of the frog kidney tissue for RT-PCR analysis were from lateral upper pole of the frog kidney which contained proximal tubules. So far, there were no similar findings in the frog kidney, but many scientists find KCNQ1 channels in rat and mouse kidney.
Previous immunohistochemistry experiments performed in the mouse kidney, using the KNCE1 knockout (-/-) mice and wild-type (+/+) mice, show that mice of both phenotypes have KCNQ1 K+ channel expression on proximal tubular epithelial cells. Only KNCE1 -/- mice had defect in proximal tubule reabsorption of glucose and electrolyte. These analyses support the notion that KCNE1 and KCNQ1 may form K+ channel complex at these sites of the tubular system.7 KCNE1 and KCNQ1 K+ channel have been localized to the brush border of the mid to late proximal tubule in mouse, where they maintain the proximal tubule brush border membrane potential, and the loss of KCNQ1/KCNE1 lead to urinary loss of Na+ and glucose.2 Compared with the wild-type mice, the addition of 5 mmol/L glucose to the luminal fluid in isolated perfused proximal tubule of KCNQ1 deficient (-/-) mice led to an enhanced rapid depolarization of the basolateral membrane cell potential as a consequence of depolarization of the luminal cell membrane.12 Consistent with the lowering of the electrochemical driving force for Na+ reabsorption in KCNQ1 -/- mice, nephrons perfused from midproximal to distal tubule with perfusate containing 5 mmol/L glucose, revealed lower absorption of Na+, fluid and glucose in the perfused nephron segment in KCNQ1 -/- compared with the wild-type mice.12 Further-more, proximal tubular Na+ reabsorption was reduced during increased luminal glucose load in response to pharmacological inhibition of KCNQ1 with its blocker HMR 1556 in the rat.12 This indicates that KCNQ1-dependent K+ movement from the cell to the lumen contributes to the maintenance of the electrical driving force for K+- coupled transport in proximal tubule. Non expression of KCNQ1 K+ channel on proximal tubules in frog kidney in the present study confirms our previous results of absence of this K+ channel protein13 on proximal epithelial cells in the same animal.
The absence of KCNQ1-dependent K+ channels in proximal tubules in this study, correspond to in situ hybridization studies on foetal and adult mouse, where KCNQ1 was expressed in the collecting ducts of the outer medulla and in the distal and collecting tubules of the cortex.8 Also, using Western analysis, Northern analysis and RT-PCR14 have confirmed expression of KCNQ1 in mouse kidney. However, they used specific antigens for particular cells to determine individual tubular segments. So they concluded that immunohistochemical localization revealed heterogeneous distribution of KCNQ1 expression, not only with respect to cell type, but also in its subcellular distribution.14 In their study they found no evidence for KCNQ1 expression in proximal tubules of mice by immunohistochemistry. Similarly to our present study in frog kidney, they found intense and basolateral KCNQ1 immunolabel in distal convoluted tubules, while in medullar collecting ducts intercalated cells showed diffuse cytoplasmatic staining. The antibodies used in our experiments correspond to the carboxy terminus of KCNQ1 protein, as did the probe used for in situ hybridization and immunohistochemistry,8,14 which also demonstrated KCNQ1 signal in distal tubules and collecting ducts in the mouse kidney.
Experiments with polarized Madin-Darby canine kidney (MDCK) cells, which share many properties of distal tubule segments, have shown a basolateral localization of KCNQ1 K+ channels, primarily determined by a tyrosine signal (Y51) in the NH2 terminal. Co-expression of KCNE+β-subunits does not alter the targeting in MDCK cells.16 The truncation mutants revealed that residues between amino acid 20 and 40 in the N-terminal tail of KCNQ1 are important for the steady-state localization to the basolateral plasma membrane.16 This statement could be the explanation for basolateral expression of KCNQ1 K+ channel in distal convoluted tubules.
The function of KCNQ1 K+ channels in the kidney has not been finally established, but KCNQ1 knockout mice were found to suffer from lower potassium sera level, urinary and faecal salt wasting, and volume depletion, thereby indicating an important role for the IsK channel in renal function.12 Basolateral K+ channels are responsible for generating the basolateral membrane potential in distal nephron and mediate K+ recycling coupled to K+ entry into cells by the Na+-K+-ATPase and influences Na+ reabsorption by helping to establish the apical membrane potential.17 Thus, depending on cellular distribution of KCNQ1 K+ channel in distal nephron, it may participate in K+ secretion and/or absorption.
Although there are marked quantitative differences, qualitatively the transport mechanisms appear to be similar in amphibian and mammalian nephrons. Direct investigations on both amphibian and mammalian nephrons have shown that the distal tubule and the collecting ducts determine the rate of urinary potassium excretion. As amphibian nephron lacks a loop of Henle, a site of significant sodium, chloride and potassium reabsorption in the mammalian nephron, some 75% of the filtered potassium load is reabsorbed along the distal tubule in amphibians. The amphibian distal tubule also has the capacity of altering magnitude and direction of potassium transfer in response to metabolic stimuli.18 Assuming the late distal tubule of amphibia to be phylogenetically equivalent to the distal convoluted tubule of a mammalian species, detailed studies of the different cell types in late distal and collecting tubule have yet to be made in non-mammalian vertebrates.19 Although the kidneys of amphibians posseses a diluting segment (early distal tubule) having virtually the same transport properties as the thick ascending limb of Henle of mammalian kidneys, amphibian kidneys can not form hyperosmotic urine because of their lack of architectural structure.20 With the use of isolated cells from the frog diluting segment three types of K+ channels have been found, but their membrane location is not known. Aldosterone which raises the cell pH by stimulating luminal Na+/H+ exchange in frog diluting segment activates one of them.21 In amphibian late distal tubules H+ secretion is accomplished by Na+/H+ exchange rather than of H+ pump.22
Whereas principal cells of rat collecting tubule are responsible for K+ secretion and Na+ reabsorption,23,24 intercalated cells of rabbit collecting duct reabsorb K+ and secrete H+ by mediation of an ATP-dependent K+/H+ exchanger accompanied by apical K+ recycling.25 As detailed studies of the different cell types in late distal and collecting tubule have yet to be made in frog, basolateral K+ channels in distal tubules of frog can facilitate Na+ entry across the luminal membrane by maintaning the normal inside-negative electrical potential of the cell, while possible role of luminal K+ channels in intercalated cells is uncertain. Whether KCNQ1 participates in these processes in the distal tubule and collecting duct remains to be determined.
In summary, KCNQ1 K+ channel is expressed in a different way in the distal tubules and collecting ducts of frog kidney. All epithelial cells of distal convoluted tubules revealed basolateral expression of KCNQ1 K+ channel, while only single cells of collecting duct, probably intercalated cells, showed diffuse cell surface staining. How KCNQ1 K+ channel contributes to epithelial transport in distinct segments of renal tubules remains to be determined.
1. Bleich M, Shan Qi-X. Epithelial K+ channels: driving force generation and K+ recycling for epithelial transport with physiological and clinical implications. Acta Physiologica Sinica 2007;59:443-53.[PubMed]
2. Jespersen T, Grunnet M, Olesen SP. The KCNQ1 potassium channel: from gene to physiological function. Physiology 2005;20: 408-16.[PubMed]
3. Lang F, Vallon V, Knipper M, Wangemann P. Functional significance of channels and transporters expressed in the inner ear and kidney. Am J Physiol 2007;293:1187-208.[PubMed]
4. Robbins J. KCNQ potassium channels: physiology, pathophysiology, and pharmacology. Pharmacol Therapeutics 2001;90:1-19.[PubMed]
5. Bleich M, Warth R. The very small-conductance K channel KvLQT1 and epithelial function. Pflugers Arch 2000;440:202-6.[PubMed]
6. Sugimoto T, Tanabe Y, Shigemoto R et al. Immunohistochemical study of a rat membrane protein which induces a selective potassium permeation: its localization in the apical membrane portion of epithelial cells. J Membr Biol 1990;113:39-47.[PubMed]
7. Vallon V, Grahammer F, Richter K et al. Role of KCNE1-dependent K+ fluxes in mouse proximal tubule. J Am Soc Nephrol 2001;12:2003-11.[PubMed]
8. Demolombe S, Franco D, Boer P et al. Differential expression of KvLQT1 and its regulator IsK in mouse epithelia. Am J Physiol Cell Physiol 2001;280:359-72.[PubMed]
9. Barhanin J, Lesage F, Gullemare E et al. K(V)LQT1 and IsK (miinK) proteins associate to form (Iks) cardiac potassium current. Nature 1996;384:78-80.
10. Sanguinetti MC, Curran ME, Zou A et al. Coassembly of K(V)LQT1 and minK (IsK) proteins to form cardiac I(Ks) potassium channel. Nature 1996;384:80-3.[PubMed]
11. Morokuma J, Blackiston D, Levin M. KCNQ1 and KCNE1 K+ channel components are involved in early left-right patterning in Xenopus laevis embryos. Cell Physiol Biochem. 2008;21:357-72.[PubMed]
12. Vallon V, Grahammer F, Volkl H et al. KCNQ1-dependent transport in renal and gastrointestinal epithelia. Proc Natl Acad Sci USA 2005;102:17864-9.[PubMed]
13. Čemerikić D, Nešović-Ostojić J, Popadić D et al. Absence of KCNQ1-dependent K+ fluxes in proximal tubular cells of frog kidney. Comp Biochem Physiol A Mol Integr Physiol 2007;148:635-44.[PubMed]
14. Zheng W, Verlander JW, Lynch J et al. Cellular distribution of the potassium channel KCNQ1 in normal mouse kidney. Am J Physiol 2007;292:456-66.[PubMed]
15. Müller CA, Markovic-Lipkovski J, Klatt T et al. Human alpha-defensins HNPs-1, -2, and -3 in renal cell carcinoma: influences on tumor cell proliferation. Am J Pathol 2002;160:1311-24.[PubMed]
16. Jespersen T, Rasmussen HB, Grunnet M et al. Basolateral localization of KCNQ1 potassium channels in MDCK cells: molecular identification of an N-terminal targeting motif. J Cell Sci 2004;117:4517-26.[FullText]
17. Gray DA, Frindt G, Zhang YY, Palmer LG. Basolateral K+ conductance in principal cells of rat CCD. Am J Physiol Renal Physiol 2005;288:493-504.[PubMed]
18. Giebisch G. Some transport properties of amphibian and mammalian nephrons. In: Comparative Physiology. Eds.: L. Bolis, K. Schmidt-Nielsen and S.H.P. Maddrell. North-Holland Publishing Co., 1973;241-91.
19. Dantzler WH. Regulation of renal proximal and distal tubule transport: sodium, chloride and organic anions. Comp Biochem Physiol A Mol Integr Physiol 2003;136:453-78.[PubMed]
20. Nishimura H., Fan Z. Regulation of water movement across vertebrate renal tubules. Comp Biochem Physiol 2003;136:479-98.[PubMed]
21. Hebert SC, Desir G, Giebisch G, Wang W. Molecular diversity and regulation of renal potassium channels. Physiol Rev 2005;85:319-71.[PubMed]
22. Lang F, Rehwald W. Potassium channels in renal epithelial transport regulation. Physiol Rev 1992;72:1-32.[PubMed]
23. Palmer L.G., Frindt G. Amiloride-sensitive Na channels from the apical membrane of the cortical collecting tubule. Proc Nat Acad Sci USA 1986;83:2767-70.[PubMed]
24. Palmer L.G. Potassium secretion and the regulation of distal nephron K channels. Am J Physiol 1999;277:821-5.[PubMed]
25. Wingo CS., Smolke AJ. Function and structure of H-K-ATPase in the kidney. Am J Physiol 1995;269:1-16.[PubMed]
[TOP]