L.M. Carrillo,1 E. Arciniegas,1 H. Rojas,2 R. Ramírez1
1Servicio Autónomo Instituto de Biomedicina, Facultad de Medicina, Universidad Central de Venezuela, Caracas; 2Instituto de Inmunología, Facultad de Medicina, Universidad Central de Venezuela, Caracas, Venezuela
Correspondence: Dr. Enrique Arciniegas, Servicio Autónomo Instituto de Biomedicina, Facultad de Medicina, Universidad Central de Venezuela, Caracas 1010, Venezuela.
Tel. +58.212.6054050 - Fax: +58.212.8611258.
E-mail: earciniegasbeta@yahoo.com
Key words: endocan, chicken embryo, aorta, focal complexes, EGF-like domain.
Acknowledgments: we thank Eng. Antonio Salgado and Mr. Wilman Clark for the photographic contribution (Area de Informática, SAIB). This study was supported by Consejo de Desarrollo Científico y Humanístico, Universidad Central de Venezuela, Caracas (Grant no. PI-09-7319-2008).
Received for publication: 22 November 2010.
Accepted for publication: 7 March 2011.
This work is licensed under a Creative Commons Attribution 3.0 License (by-nc 3.0).
©Copyright L.M. Carrillo et al., 2011
Licensee PAGEPress, Italy
European Journal of Histochemistry 2011; 55:e13
doi:10.4081/ejh.2011.e13
AbstractEndocan is a dermatan sulfate proteoglycan (DSPG) that has been observed in the cytoplasm of endothelial cells of small and large vessels in lung, kidney, liver, colon, ovary and brain tumors. This DSPG has been implicated in the regulation of cellular activities such as adhesion, migration, and proliferation. Given the important roles played by endocan in such processes, we sought to determine whether this DSPG is present in the chicken embryo aortic wall in embryonic days 12 and 14, when intimal thickening and endothelial transformation are notorious. Immunolabeling of serial paraffin cross-sections revealed endocan immunoreactivity at the endothelium and some mesenchymal cells constituting the intimal thickening but not in the cells arranged in lamellar layers. We also investigated whether endocan was present in monolayers of primary embryonic aortic endothelial cells attached to fibronectin when they were deprived of serum and stimulated with epidermal growth factor. Immunofluorescence determined that in the epidermal growth factor (EGF) condition where separating, detaching, and migrating cells were observed, endocan appeared organized in arrays typical of focal complexes in the leading edge of these cells. In serum-free medium condition in which the endothelial cells displayed a cobblestone appearance, endocan appeared mainly delineating the margin of many cells. This study demonstrates for the first time the presence of endocan during the aortic wall remodeling, and provides evidence that suggests a possible contribution of this DSPG in the endothelial-mesenchymal transition (EndoMT) process. |
Endothelial-mesenchymal transition (EndoMT) is a complex process by which certain endothelial cell subsets lose endothelial characteristics and transform into mesenchymal or smooth muscle cells (SMCs).1 This process is now recognized as a phenomenon that occurs during cardiac fibrosis2 and neointimal thickening formation observed in atherosclerosis and restenosis.1,3 In addition to this role, recent evidence has also demonstrated that EndoMT is critical in heart and vascular development,1,4 hypertrophic scarring,5 kidney fibrosis,6 diabetic nephropathy7 and during cancer progression.8 EndoMT process entails growth factor and cytokines synthesis and secretion, activation of receptor tyrosine kinases (RTKs) and non-RTKs, cytoskeleton reorganization, disassembly of the adherence junctions or loss of endothelial cell-cell contacts, change in cell shape and polarity, proteases secretion, extracellular matrix (ECM) remodeling, cell separation or detachment from the substratum, and cell migration and differentiation.1,9
In addition to permanent interaction with other cells, endothelial cells also need to interact with the ECM. It is now well established that most ECM molecules participate in tissue remodeling by promoting cell adhesion provoking cytoskeletal organization, and regulating cell spreading, polarization, detachment, migration and differentiation in response to mechanical stimuli and/or cross talk of a variety of growth factors and cytokines.10 Some of the most studied ECM molecules in the vessel wall belong to the family of proteoglycans (PGs), which are actually recognized by its role in several cellular processes either via interactions with ECM receptors like integrins or via signaling through receptor tyrosine kinases (RTKs).11 Some particularly significant studies on the developing of chick aorta show that large PGs containing EGF-like domains such as versican and aggrecan have different spatial and temporal expression patterns, suggesting a possible role for these molecules during intimal thickening formation and EndoMT.11,12
Endocan, also called endothelial cell-specific molecule-1, is a small dermatan sulfate proteoglycan (DSPG) that does not seem to belong to the small leucine-rich PGs (SLRPG) family and is mainly expressed by endothelial cells and regulated in vivo by inflammatory cytokines such as vascular endothelial growth factor (VEGF), fibroblast growth factor-2 (FGF-2), and tumor necrosis factor-α (TNF-α).13,14,15,16 This DSPG consist of a core protein that contains an N-terminal rich in cysteine residues, a putative EGF-like domain, whose functional significance is not yet fully understood, and a single chain of dermatan sulfate (DS), which is covalently linked to the core protein.15 Interestingly, a marked expression of endocan has been observed in breast, brain, lung, liver, kidney, ovary tumors, thus suggesting an important role of endocan in angiogenesis and in the development and progression of cancer.16 Nevertheless, the factors regulating the synthesis and secretion as well as their functions, have not yet been fully elucidated.
In view of the above observations, the aim of this study was to investigate immunohistochemically whether endocan is present in the chicken embryo aortic wall in embryonic days 12 and 14 (days E12 and E14; stages 38 and 40), when intimal thickening and endothelial transformation are notorious. We also investigated the presence of endocan in monolayers of primary embryonic aortic endothelial cells attached to fibronectin (FN) and stimulated with epidermal growth factor (EGF).
Fertilized chicken eggs (White leghorn) were obtained from a local hatchery (Granja Avícola Agropollito C.A., Paracotos, Estado Miranda, Venezuela) and incubated at 37ºC and 60% humidity for 12 and 14 days (stages 38 and 40). Embryos were staged according to Hamburger and Hamilton.17 The aortae were dissected were placed in Hank’s balanced salt solution without Ca++ and Mg++ (HBSS) (Sigma-Aldrich, St. Louis, MO, USA), and fixed for 15 min at room temperature with 4% formaldehyde prepared from paraformaldehyde in phosphate-buffered saline (PBS). The aortae were dehydrated in graded ethanol and embedded in paraffin. Paraffin sections (5 μm thick) were mounted on silanized slides (Dako, Inc., Carpinteria, Ca, USA). A total of two aortae for each stage obtained from three different lots of fertilized chicken eggs were processed.
Serial deparaffinized cross sections were air dried and equilibrated in PBS for 10 min. Non-specific antibody staining was blocked by incubating sections in PBS containing 2% bovine serum albumin (BSA) for 30 min at room temperature. Sections were incubated overnight at 4ºC in a humid chamber with mouse monoclonal anti-human endocan/ESM-1 antibody (clone MEP08) (Lunginnov, Lille, France) or mouse monoclonal antibody anti α-SM actin (clone 1A4) (Sigma-Aldrich). After the sections were washed several times in PBS, anti-mouse FITC-conjugated secondary antibody (Dako) was applied for 30 min. The sections were again washed in PBS, and coverslipped with mounting medium (IMMU-mount; Shandon, Pittsburg, PA, USA). Negative controls were produced by the use of purified normal serum or PBS in place of primary antibody. A IX81 Olympus inverted microscope with the Fluo-View confocal laser scanning configuration (CLSM) (Olympus America, Inc., Melville, NY, USA) was used to examine the sections. Fluorescence intensity was measured by using a processing software program (FV10. ASW version 02.01.01.04, Olympus Corporation).
Aortae from days E11-E12 (stages 37 and 38) were dissected in HBSS (Sigma-Aldrich) at 37ºC. Segments, approximately 8 mm2 in surface area, were isolated (distal to the aortic arches) and opened along longitudinal axis. Explants were rinsed in HBSS (Sigma-Aldrich) and left in medium 199 with Earle’s salts with L-Glutamine (Sigma-Aldrich) containing 0.025% BSA (Sigma-Aldrich), 100 μg/mL streptomycin, and 100 U/mL penicillin (GIBCO, Invitrogen, Carlsbad, CA, USA) for 5-10 min before starting the assays. For simplicity, this medium is referred to as serum-free medium (SFM). Aortic explants were placed with the endothelial apical side down on fibronectin-coated 35 mm Petri dishes (Nunclon, Delta, IL, USA) containing 300 μL of SFM. They were incubated at 37ºC in a humidified atmosphere consisting of 5% C02 and 95% air for 4 h. After this interval, 1 mL of SFM was gently added to each dish. Two h later, the adhered explants were removed with the aid of a thin needle, leaving a monolayer of retracted endothelial cells that exhibited zones denuded of cells or wounds. Monolayers were examined with an inverted microscope (IX-70 Olympus, Olympus America Inc.). Cell cultures were initiated when the monolayers of endothelial cells were rinsed 5 times with HBSS and incubated for 2 h at 37ºC with 5% CO2. At the end of this time, the medium of the monolayers was switched to SFM supplemented with recombinant epidermal growth factor (EGF) (R&D Systems, Inc., Minneapolis, MN, USA) (100 ηg/mL) and 0.1% chicken serum (ChS) (GIBCO) and incubated for an additional 10-12 h period.
Fixed and permeabilized cells were processed for immunostaining as described above using the following antibodies: a mouse monoclonal anti-human endocan/ESM-1 (clone MEP08) (Lunginnov), a rabbit polyclonal anti-human von Willebrand´s factor (vWf) (Dako), a mouse monoclonal antibody raised against native chick brain microtubules (clone DM1A) (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), a mouse monoclonal antibody anti-human β3-integrin (GPIIIa, CD61) (clone 25E11) (Millipore Chemicon Corporation, Billerica, MA, USA), and a rabbit polyclonal anti-phospho-FAK (Tyr-397) (Sigma-Aldrich). Negative controls were performed by omitting the primary antibody incubation step or by using non-immune serum instead of primary antibody. A confocal laser scanning microscope (CLSM) (Eclipse TE-300 Nikon inverted microscope) (Nikon Instruments Inc., Melville, NY, USA) equipped with a Nikon objective Plan-Apo BC x60, 1.2 wi coupled to a C1-LU2 unit Argon cooled air (488 ηm) laser was used to examine the cultures. The laser unit was controlled by a D-eclipse C1 interface.
In order to investigate the expression and localization of endocan in the aortic wall at days E12 and E14 (stages 38 and 40) of development when intimal thickening is distinguishable and EndoMT occurs,18 serial paraffin sections were examined by confocal microscopy. A specific monoclonal antibody that recognizes epitopes on the N-terminus of endocan was used.
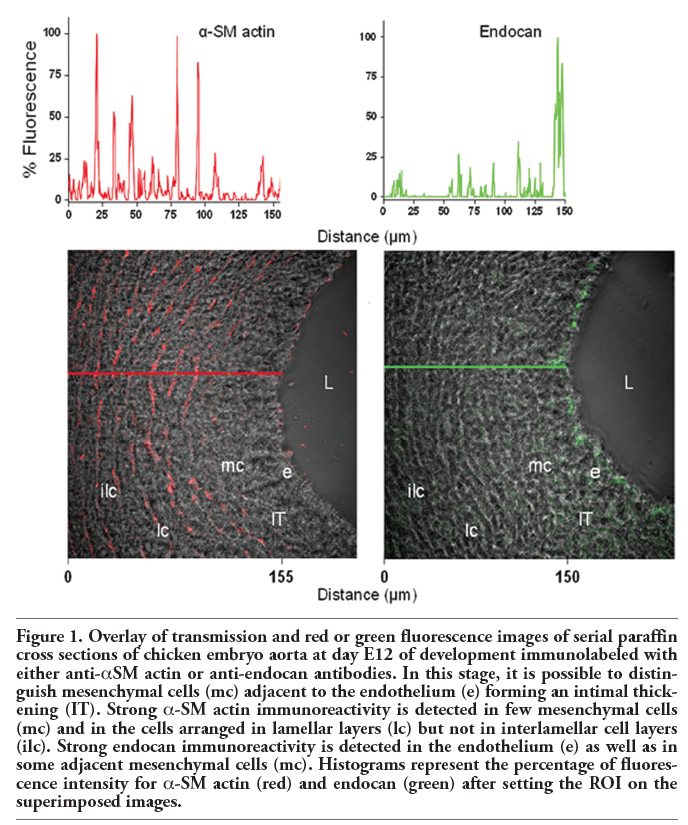
At 12-14 days of development, the aortic wall is composed by the endothelium, which limits the vessel lumen, and radially oriented mesenchymal cells originating from the endothelium and that constitute the intimal thickening. At these stages it is also possible to distinguish cells organized into circular lamellar and interlamellar layers that form the media. Occasionally, some mesenchymal cells and lamellar cell layers are observed displaying immunoreactivity for α-smooth muscle actin (α-SM actin) (Figure 1). At these stages, a strong immunoreactivity for endocan was found at the endothelium and some mesenchymal cells constituting the intimal thickening but not in the cells arranged in lamellar layers (Figure 1).
No immunoreactivity was detected when the primary antibody was omitted or replaced by non-immune serum in control sections (not shown).
|
Figure 1. Overlay of transmission and red or green fluorescence images of serial paraffin cross sections of chicken embryo aorta at day E12 of development immunolabeled with either anti-αSM actin or anti-endocan antibodies. In this stage, it is possible to distinguish mesenchymal cells (mc) adjacent to the endothelium (e) forming an intimal thickening (IT). Strong α-SM actin immunoreactivity is detected in few mesenchymal cells (mc) and in the cells arranged in lamellar layers (lc) but not in interlamellar cell layers (ilc). Strong endocan immunoreactivity is detected in the endothelium (e) as well as in some adjacent mesenchymal cells (mc). Histograms represent the percentage of fluorescence intensity for α-SM actin (red) and endocan (green) after setting the ROI on the superimposed images. |
Based on our findings in vivo, we also investigated whether endocan was present in monolayers of embryonic endothelial cells attached to FN that had been transiently mechanically altered during the explant removal, when they were deprived of serum for 2 h and switched to medium supplemented with EGF and ChS and maintained for an additional 10 or 12 h period.
Immunofluorescence analyzed by confocal microscopy determined that in the EGF condition in which separating, detaching, and migrating endothelial cells are observed and the vWf expression is notorious (Figure 2 a,b), all of the cells displayed strong immunoreactivity for endocan (Figure 2 c,d). Specifically, this DSPG in addition to being located delineating the margins of some endothelial cells, was also located in punctate and linear arrays typical of focal complexes at the leading edge as well as in the area surrounding the microtubule-organizing center (MTOC) and aligned with microtubules (MTs) of separating, detaching, and migrating cells (compare panels c and e in Figure 2). Focal adhesion complexes are considered not only as structural and dynamic links between the ECM and the cytoskeleton controlling cell shape, spreading and motility, but also as sites for signal transduction through integrins and adaptor proteins such as focal adhesion kinase (FAK) and paxillin that are present in these sites. We therefore studied the presence, organization and distribution of these proteins by confocal microscopy in monolayers of embryonic endothelial cells attached to FN and stimulated with EGF for 10-12 h. In such condition, immunofluorescence staining with anti-integrin-β3 and anti-p-FAK revealed that many cells displayed immunoreactivity with a location and distribution resembling to that observed for endocan. Specifically, integrin-β3 and p-FAK appeared delineating the margins of some cells and organized into linear streaks and a punctate pattern at the leading edge of the cells that were separating, detaching, and migrating (Figure 3 a,b). In the SFM condition in which the endothelial cells displayed a cobblestone appearance and neither separating, detaching nor migrating cells were observed, endocan, integrin-β3 and p-FAK appeared mainly distributed delineating the margins of many cells (Figure 4 a,b,c).
No immunolabeling was observed when a non-immune serum was used as negative control (not shown).
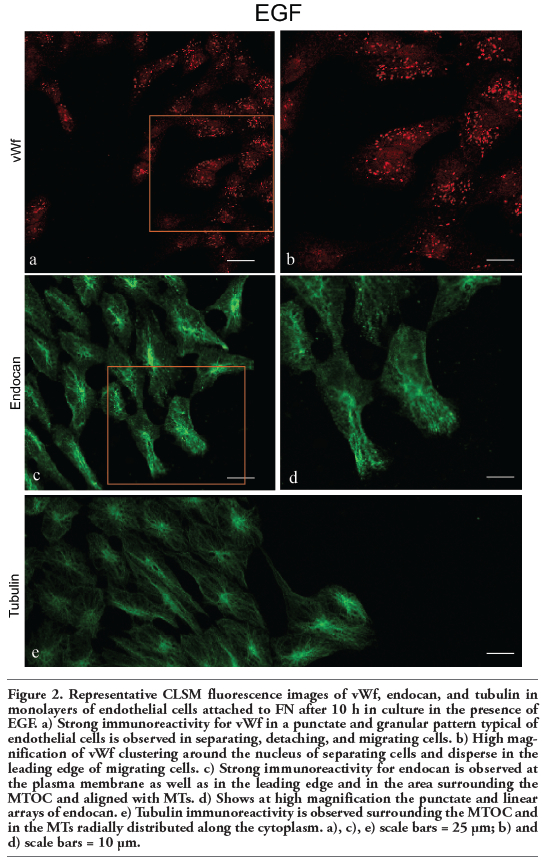 |
Figure 2. Representative CLSM fluorescence images of vWf, endocan, and tubulin in monolayers of endothelial cells attached to FN after 10 h in culture in the presence of EGF. a) Strong immunoreactivity for vWf in a punctate and granular pattern typical of endothelial cells is observed in separating, detaching, and migrating cells. b) High magnification of vWf clustering around the nucleus of separating cells and disperse in the leading edge of migrating cells. c) Strong immunoreactivity for endocan is observed at the plasma membrane as well as in the leading edge and in the area surrounding the MTOC and aligned with MTs. d) Shows at high magnification the punctate and linear arrays of endocan. e) Tubulin immunoreactivity is observed surrounding the MTOC and in the MTs radially distributed along the cytoplasm. a), c), e) scale bars = 25 μm; b) and d) scale bars = 10 μm. |
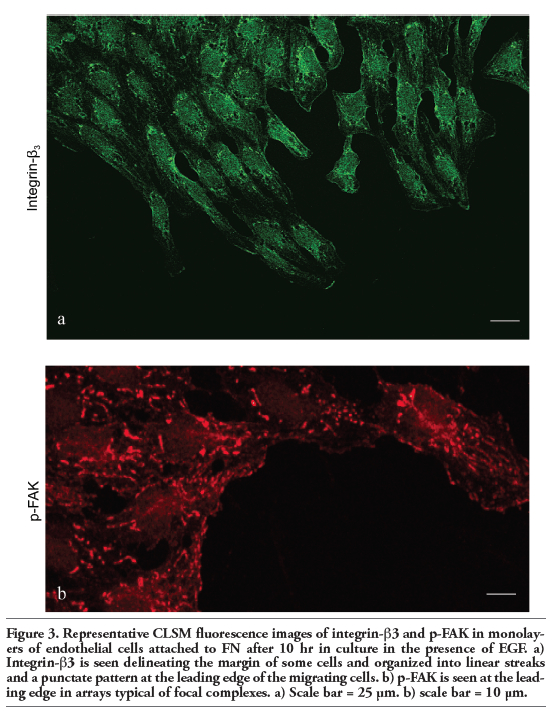 |
Figure 3. Representative CLSM fluorescence images of integrin-β3 and p-FAK in monolayers of endothelial cells attached to FN after 10 hr in culture in the presence of EGF. a) Integrin-β3 is seen delineating the margin of some cells and organized into linear streaks and a punctate pattern at the leading edge of the migrating cells. b) p-FAK is seen at the leading edge in arrays typical of focal complexes. a) Scale bar = 25 μm. b) scale bar = 10 μm. |
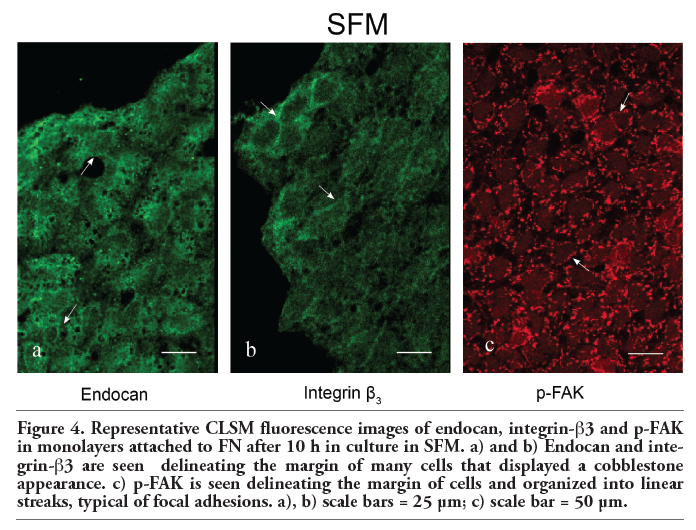 |
Figure 4. Representative CLSM fluorescence images of endocan, integrin-β3 and p-FAK in monolayers attached to FN after 10 h in culture in SFM. a) and b) Endocan and integrin-β3 are seen delineating the margin of many cells that displayed a cobblestone appearance. c) p-FAK is seen delineating the margin of cells and organized into linear streaks, typical of focal adhesions. a), b) scale bars = 25 μm; c) scale bar = 50 μm. |
The current study reveals the existence of both endocan-immunoreactive endothelial and mesenchymal cells in the intimal thickenings observed in chicken embryo aortae on days E12 and E14. This study also provides evidence that allow us to suggest a possible contribution of this DSPG in the EndoMT process.
Endocan is a DSPG expressed specifically by endothelial cells and has been implicated in several pathophysiological processes as well as in the regulation of cellular activities such as adhesion, migration and proliferation.14,16 By immunohistochemistry, endocan has been observed in the cytoplasm of endothelial cells of small and large vessels in lung, kidney, liver, colon, ovary and brain tumors.16 Thus, it is of interest that a strong endocan staining was detected in the endothelium and some migrating mesenchymal cells that conform the spontaneous intimal thickening but not in those cells organized in lamellar layers. This implies that endocan would be participating in the EndoMT and therefore contributing to the aortic wall remodeling. Importantly, spontaneous aortic and pulmonary intimal thickenings have been demonstrated during chicken embryo development and related to blood pressure alterations that occur rapid and exponentially from day E3 to day E10 of development accompanied by the local production and activation of different growth factors such as FGF-2, TGFβ and IGFII.19,20 It might thus be speculated that the presence of endocan in the embryonic aortic wall would occur in response to hemodynamic changes and production of certain growth factors, considering that endocan expression, synthesis or secretion is highly regulated by VEGF and FGF-2,14,16 and that TGFβ stimulates the synthesis and secretion of the core protein of some PGs.21,22 Also, this presence may be interpreted as a sign of endothelial activation.
In this study, we also found that the spatial distribution and organization of endocan observed in SFM condition was altered when the monolayers were stimulated with EGF. Specifically, endocan appeared delineating the margins of cells in arrays typical of focal complexes in the leading edges as well as surrounding the MTOC and aligned with MTs of the cells that were separating, detaching and migrating toward cell-free areas. We believe that this peculiar localization of endocan would be linked with a participation in the assembly, disassembly, and signaling of focal adhesions, in which integrin-β3 clustering induced by FN and activation of adaptor proteins, including focal adhesion kinase (FAK) and paxillin that interact with cytoskeletal proteins such as microtubules and actin microfilaments, are required.23,24,25,26,27 Moreover, considering that inmunoreactivities observed for integrin-β3 and p-FAK in separating, detaching and migrating cells, resembled to that observed for endocan.
As suggested, the function of DS-chain of endocan is to bind some growth factors and cytokines as well as ECM molecules including FN and collagen, but the functional significance of its EGF-like is not yet fully understood.15 It is likely that endocan, through its EGF-like domain, directly binds to integrin-β3, a cell surface receptor that is present on the apical side of endothelial cells,28,29 and that this binding occurs in the presence of divalent cations, considering that endocan interacts physically and functionally with the integrin lymphocyte function associated-antigen-1 (LFA-1), present in leukocyte and Jurkat cells, through its polypeptide moiety13 and that the binding of growth factors containing EGF-like motifs to integrin α5β3 occurs via its EGF-like motif.30 Also, it is possible that endocan through its DS-chain binds to FN, facilitating the endothelial cell adhesion. Based on these asseverations and our findings, we hypothesized that endocan might be cooperating with integrins to promote focal complexes assembly and disassembly and hence influencing cell separation, detachment and migration, important steps for the EndoMT.
1. Arciniegas E, Frid MG, Douglas IS, Stenmark KR. Perspectives on endothelial-to-mesenchymal transition: potential contribution to vascular remodeling in chronic pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol 2007;293:L1-L8.[PubMed]
2. Goumans MJ, van Zonneveld AJ, ten Dijke T. Transforming growth factor β-induced endothelial-to-mesenchymal transition: A switch to cardiac fibrosis? Trends Cardiovasc Med 2008;18:293-8.[PubMed]
3. Morrell NW, Adnot S, Archer SL, Dupuis J, Jones PL, MacLean MR, et al. Cellular and molecular basis of pulmonary arterial hypertension. J Am Coll Cardiol 2009;54:S20-31.[PubMed]
4. Mironov V, Visconti RP, Markwald RR. On the role of shear stress in cardiogenesis. Endothelium 2005;12:259-61.[PubMed]
5. Xi-Qiao W, Ying-Kai L, Chun Q, Shu-Liang L. Hyperactivity of fibroblasts and functional regression of endothelial cells contribute to microvessel occlusion in hypertrophic scarring. Microvasc Res 2009;77:204-11.[PubMed]
6. Zeisberg EM, Potenta SE, Sugimoto H, Zeisberg M, Kalluri R. Fibroblasts in kidney fibrosis emerge via endothelial-to-mesenchymal transition. J Am Soc Nephrol 2008;19:2282-7.[PubMed]
7. Li J, Bertram J. Review: Endothelial-myofibroblast transition, a new player in diabetic renal fibrosis. Nephrology 2010;15:507-12.[PubMed]
8. Potenta S, Zeisberg E, Kalluri R. The role of endothelial-to-mesenchymal transition in cancer progression. Br J Cancer 2008;99:1375-9.[PubMed]
9. Baum B, Settleman J, Quinlan MP. Transitions between epithelial and mesenchymal states in development and disease. Semin Cell Dev Biol 2008;19:294-308.[PubMed]
10. Mammoto T, Ingber DE. Mechanical control of tissue and organ development. Development 2010;137:1407-20.[PubMed]
11. Wagenseil JA, Mecham RP. Vascular extracellular matrix and arterial mechanics. Physiol Rev 2009;89:957-89.[PubMed]
12. Arciniegas E, Neves CY, Candelle D, Parada D. Differential versican isoforms and aggrecan expression in the chicken embryo aorta. Anat Rec A Discov Mol Cell Evol Biol. 2004;279:592-600.[PubMed]
13. Béchard D, Scherpereel A, Hammad H, Gentina T, Tsicopoulos A, Aumercier M, et al. Human endothelial-cell specific molecule-1 binds directly to the integrin CD11a/CD18 (LFA-1) and blocks binding to intercellular adhesion molecule-1. J Immunol 2001;167:3099-106.[PubMed]
14. Abid MR, Yi X, Yano K, Shih SC, Aird WC. Vascular endocan is preferentially expressed in tumor endothelium. Microvasc Res 2006;72:136-45.[PubMed]
15. Sarrazin S, Adam E, Lyon M, Depontieu F, Motte V, Landolfi C, et al. Endocan or endothelial cell specific molecule-1 (ESM-1): a potential novel endothelial cell marker and a new target for cancer therapy. Biochim Biophys Acta 2006;1765:25-37.[PubMed]
16. Sarrazin S, Maurage CA, Delmas D, Lassalle P, Delehedde M. Endocan as a biomarker of endothelial dysfunction in cancer. J Cancer Sci Ther 2010;2:047-052.[FullText]
17. Hamburger V, Hamilton HL. A series of normal stages in the development of the chick embryo. Dev Dyn 1992;195:231-72.[PubMed]
18. Arciniegas E, Ponce L, Hartt Y, Graterol A, Carlini RG. Intimal thickening involves transdifferentiation of embryonic endothelial cells. Anat Rec 2000;258:47-57.[PubMed]
19. Arciniegas E, Parada D, Graterol A. Mechanically alterated embryonic chicken endothelial cells change their phenotype to an epithelioid phenotype. Anat Rec A Discov Mol Cell Evol Biol 2003;270:67-81.[PubMed]
20. Arciniegas E, Neves YC, Carrillo LM. Potential role for insulin-like growth factor II and vitronectin in the endothelial-mesenchymal transition process. Differentiation 2006;74:277-92.[PubMed]
21. Bassols A, Massagué J. Transforming growth factor beta regulates the expression and structure of extracellular matrix chondroitin/dermatan sulfate proteoglycans. J Biol Chem 1988;263:3039-45.[PubMed]
22. Schönherr E, Järveläinen HT, Sandell LJ, Wight TN. Effects of platelet-derived growth factor and transforming growth factor-beta 1 on the synthesis of a large versican-like chondroitin sulfate proteoglycan by arterial smooth muscle cells. J Biol Chem 1991;266:17640-7.[PubMed]
23. Adair BD, Xiong JP, Maddock C, Goodman SL, Arnaout MA, Yeager M. Three-dimensional EM structure of the ectodomain of integrin αvβ3 in a complex with fibronectin. J Cell Biol 2005;168:1109-18.[FullText]
24. Mitra SK, Hanson DA, Schlaepfer DD. Focal adhesion kinase: in command and control of cell motility. Nat Rev Mol Cell Biol 2005;6:56-68.[PubMed]
25. Romer LH, Birukov KG, Garcia JG. Focal adhesions. Paradigm for a signaling nexus. Circ Res 2006;98:606-16.[PubMed]
26. Efimov A, Kaverina I. Significance of microtubule catastrophes at focal adhesions sites. Cell Adh Migr 2009;3:285-7.[PubMed]
27. Desgrosellier JS, Cheresh DA. Integrins in cancer: biological implications and therapeutic opportunities. Nat Rev Cancer 2010; 10:9-22.[PubMed]
28. Bombeli T, Schwartz BR, Harlan JM. Adhesion of activated platelets to endothelial cells: Evidence for GPIIbIIIa-dependent bridging Mechanism and novel roles for endothelial intercellular adhesion molecule 1 (ICAM-1), αvβ3 integrin, and GPIbα. J Exp Med 1998;187:329-39.[PubMed]
29. Hauschner H, Landau M, Seligsohn U, Rosenberg N. A unique interaction between αIIb and β3 in the head region is essential for outside-in signaling-related functions of αIIbβ3 integrin. Blood 2010;115:4542-50.[Abstract]
30. Leguchi K, Fujita M, Ma Z, Davari P, Taniguchi Y, Sekiguchi K, et al. Direct binding of the EGF-like domain of neuregulin-1 to integrins (αvβ3 and α6β4) is involved in neuregulin-1/ErbB signaling. J Biol Chem 2010;285:31388-98. [Abstract]
[TOP]