
T. Nara,1 T. Yasui,1 O. Fujimori,2 W. Meyer,3 A. Tsukise1
1Department of Veterinary Anatomy, College of Bioresource Sciences, Nihon University, Kanagawa, Japan; 2Laboratory of Anatomy and Histochemistry, Faculty of Rehabilitation Science, Nagoya Gakuin University, Aichi, Japan; 3Institute for Anatomy, University of Veterinary Medicine Hannover Foundation, Hannover, Germany
Correspondence: Prof. Dr. Azuma Tsukise, Department of Veterinary Anatomy, College of Bioresource Sciences, Nihon University, 1866 Kameino, Fujisawa, Kanagawa 252-0880, Japan. Tel. +81.466.843629 - Fax: +81.466.843629.
E-mail: tsukise@brs.nihon-u.ac.jp
Key words: anal glands, sialic acids, antimicrobial substances, histochemistry, dog.
Received for publication: 5 April 2011.
Accepted for publication: 27 June 2011.
This work is licensed under a Creative Commons Attribution NonCommercial 3.0 License (CC BY-NC 3.0).
©Copyright T. Nara et al., 2011
Licensee PAGEPress, Italy
European Journal of Histochemistry 2011; 55:e29
doi:10.4081/ejh.2011.e29
AbstractThe functional properties of sialic acids appear to be manifold. Additionally, antimicrobial substances serve as a non-specific defense against microorganisms. In this study, therefore, the localization of sialic acids and antimicrobial substances in the anal glands of dog was studied by sialoglycoconjugate histochemistry and immunohistochemistry. The secretory epithelium, luminal secretions and excretory ducts exhibited high levels of sialoglycoconjugates that terminated in Siaα2-6Gal/GalNAc or Siaα2-3Gal1-4GlcNAc. Additionally, O-acetylated sialic acids were detectable in these glandular structures. Antimicrobial substances, such as lysozyme, immunoglobulin A (IgA), lactoferrin and the peptide group of β-defensins, were also demonstrated as products of the anal glands. The results obtained are discussed with regard to the functional significance of the anal glands. These secretory products may create a defensive barrier against microbial invasion at the anal mucosa. |
The canine anal region is characterized by three typical glandular complexes, which are the anal glands, circumanal glands and anal sac glands.1,2,3,4,5,6 In dog and pig, previous studies demonstrated the distribution of glycoconjugates in the anal glands using lectin histochemical methods. These glandular secretions seem to participate in the formation of a stable mucous coat of the rather dry feces and enable the marking of territory.7,8 The anal glands are also observed in human anal canal.9 Although several investigations have been reported on these glands in humans,10,11,12,13 their functional significance is mostly unexplained.
Within various saccharide residues, sialic acids in particular possess diverging functional properties. They are involved in the regulation of many cellular and molecular interactions and contribute to the high viscosity of mucus.14,15 Additionally, in mammalian integument, some antimicrobial substances are released in response to stimuli and rapidly become available at sites of microbial invasion.16,17,18 In certain domesticated mammals, secretory glycoconjugates including sialic acids and antimicrobial substances elaborated by the skin glands may have functional significance in preserving skin integrity.19,20,21,22 However, detailed localization of sialic acids and antimicrobial substances in the anal glands of dogs has not been determined.
The present study on the canine anal glands investigated the distribution of sialic acids and antimicrobial substances using glycoconjugate histochemical and immunohistochemical methods. The results obtained are believed to provide data on the specific functional properties of this gland type, not only in the dog but in other mammalian species as well.
All experiments were performed in accordance with the guidelines for the care and use of laboratory animals at the Institute of Experimental Animal Science, College of Bioresource Sciences, Nihon University. Four adult male Beagle dogs (1.5-2 years, 10-12 kg) were deeply anesthetized and then exsanguinated from the common carotid arteries. After bloodletting, the anal region was removed surgically. The tissue specimens were fixed in Bouin’s solution for 24 h at room temperature, then carefully washed and dehydrated through a graded series of ethanol. The materials were embedded in paraffin wax and then sectioned at a thickness of 5 μm. Sections were stained with hematoxylin-eosin (H-E) and Mallory-Azan as well as with sialoglycoconjugate histochemical and immunohistochemical procedures including combined control procedures.
For the localization of sialic acid, the histochemical staining method employed was periodic acid-phenylhydrazinium chloride-thiocarbohydrazide-silver proteinate-physical development (PA-P-TCH-SP-PD).23 The PA-P-TCH-SP-PD procedure is an efficient and sensitive histochemical method for demonstration of sialic acids using physical development procedure in light microscopy. Selective periodate oxidation (0.4 mM PA in 1 M HCl) of sialic acid residues oxidizes monoaldehyde at C-7 or C-8, which then condenses with phenylhydrazine to give rise to the corresponding phenylhydrazone. This compound can undergo solvolysis to yield TCH-SP-PD reactive monoaldehydes. Although the selective periodate oxidation also turns neutral sugars into the dialdehydes, they condense with phenylhydrazine to result in TCH-SP-PD unreactive azido or morpholine. On the other hand, sialic acids with O-acetyl substitutions at C-8 and/or C-9 are undetected by this procedure. Therefore, chemical modification by saponification (0.5% potassium hydroxide-70% ethanol) (KOH)24 was performed on some sections, prior to PA-P-TCH-SP-PD staining, since KOH treatment is known to remove the acetyl groups on C-4, C-7, C-8 and C-9.25 Enzyme digestion with sialidase (from Arthrobacter ureafaciens, Nacalai Tesque Inc., Kyoto, Japan; 1.0 U/mL in 0.1 M phosphate buffer, pH 5.3, at 37˚C for 18 h)26 after saponification was conducted before the PA-P-TCH-SP-PD staining. Neuraminidases from A. ureafaciens can hydrolyze α2-3, α2-6 and α2-8 linkages.27 However, O-acetylated sialic acids, especially at C-4, resist hydrolytic cleavage by bacterial neuraminidase.28 These physical development procedures for detection of sialic acids and their moieties are summarized in Table 1.29,30,31 Additionally, some sections were incubated with biotinylated Sambucus sieboldiana agglutinin (SSA) and Maackia amurensis agglutinin (MAM) (Seikagaku Kogyo Co., Tokyo, Japan) at concentrations of 10-20 μg/mL in 0.05 M phosphate-buffered saline (PBS) (pH 7.2) for 60 min at room temperature, following treatment with 0.3% hydrogen peroxide to suppress endogenous peroxidase (PO) activity and then preincubation with 1% bovine serum albumin (BSA) in PBS for 30 min at room temperature. The lectin specificities and inhibitory sugars are listed in Table 2.32 After rinsing with PBS, the sections were soaked in PO-labeled streptavidin (Nichirei Co., Tokyo, Japan) for 30 min at room temperature. The activity of the PO employed for labeling was evaluated with a solution containing 0.05 M Tris-HCl buffer (pH 7.6), diaminobenzidine (DAB) and hydrogen peroxide. Lectin controls were prepared by addition of inhibitory sugars at a final concentration of 0.01 M to the respective lectin solutions, by substitution of unconjugated lectins for biotinylated lectin conjugates or by exposure of sections to the PO-DAB system without lectin
For the immunohistochemical methods, the sections were preincubated with Proteinase K solution (Dako, Glostrup, Denmark) for 3 min at room temperature and treated with 0.3% hydrogen peroxide for 30 min for the blocking of endogenous PO activities. Then, they were incubated with primary antibodies diluted in 0.01 M PBS (pH 7.3) against IgA (polyclonal, from rabbit) (Dako), lysozyme (polyclonal, from rabbit) (Dako), lactoferrin (polyclonal, from rabbit) (MP Biomedicals, Ohio, USA) and human β-defensin-1 (polyclonal, from rabbit) (Biologo, Kronshagen, Germany) for 24 h at 4°C, following pretreatment with 1% BSA in PBS for 30 min at room temperature. The primary antibodies were diluted as follows: IgA 1:100, lysozyme 1:100, lactoferrin 1:100, human β-defensin-1 1:1000. After rinsing with PBS, these sections were incubated with prediluted dextran-polymer-conjugated secondary antibodies (EnVision+ Dual Link System-HRP, raised goat against mouse and rabbit immunoglobulins, Dako) for 30 min at room temperature. The immunoreaction employed for labeling was revealed using a DAB hydrogen peroxide system, as described above. Controls for immunohistochemical methods were performed by incubation with PBS without primary antibodies or exposure of sections to the PO-DAB system without primary or secondary antibodies.
|
Table 1. Physical development procedures for detection of sialic acids and their moieties.21,30,31 |
 |
Table 2. The lectins used and their sugar residues and inhibitory sugars. |
In the anal canal of dog, the anal glands occupied the submucosa of the columnar and intermediate zones covered with stratified squamous epithelium or outside the internal anal sphincter muscle (Figure 1a). The anal glands were tubuloalveolar glands (Figure 1b,c) and their longer excretory ducts passed through the muscle layers, and directly opened on to the epithelial surface. Additionally, the secretory portions and excretory ducts of these glands were often dilated. The PA-P-TCH-SP-PD procedure, which demonstrates the localization of sialic acid, produced moderate to weak reactions in the secretory cells and excretory duct cells, while the luminal secretions reacted very strongly (Figure 2a). Saponification clearly increased the intensity of the PA-P-TCH-SP-PD reaction (Figure 2b). The PA-P-TCH-SP-PD staining intensity was nearly negative after sialidase digestion following saponification (Figure 2c). When the sections were subjected to the lectin histochemical procedures, the SSA technique resulted in strong to moderate reactions of the secretory cells and luminal secretions (Figure 3a). A moderate reaction was observed in the excretory ducts after the application of SSA. In the sections stained for MAM, the luminal secretions and excretory duct cells showed distinct positive reactions (Figure 3b). The MAM staining patterns of the secretory cells were similar to the results obtained by the PA-P-TCH-SP-PD staining, showing a moderate to weak reaction (Figure 3c). In all of the control experiments using the lectin histochemical procedures, the positive lectin reactions were greatly diminished or nearly abolished in the glandular structures (Figure 3d). The results obtained for the sialoglycoconjugate histochemical staining are summarized in Table 3.
For the localization of antimicrobial substances, antibodies to IgA showed strong to moderate reactions in the luminal secretions and excretory ducts (Figure 4a). The luminal surface and apical cytoplasm of the secretory cells were mainly found to exhibit distinct positive reactions after treatment with antibodies to IgA (Figure 4b). Additionally, in some of the secretory cells, the cytoplasm was stained strongly. A strong positive reaction was also observed in the plasma cells, which were distributed in the connective tissues surrounding the anal glandular acini (Figure 4b). When the tissue sections were incubated with anti-lysozyme, the staining intensity differed with the secretory cells showing moderate to very weak positive reactions (Figure 4c). The luminal secretions and excretory ducts exhibited moderate to weak positive reactions. A moderate positive reaction for lactoferrin was detectable in the secretory epithelium, whereas the luminal secretions showed a weak reaction (Figure 4d). The reaction intensity of the excretory ducts was strong to moderate. Incubation with antibodies to β-defensin 1 led to the same staining images as those obtained with antibodies to lactoferrin (Figure 4e). In control experiments for immunohistochemical staining performed by incubation with PBS without primary antibodies or exposure of sections to the PO-DAB system without primary or secondary antibodies, no glandular structures exhibited any positive reactions (Figure 4f). These results obtained from the immunohistochemical staining for antimicrobial substances are summarized in Table 4.
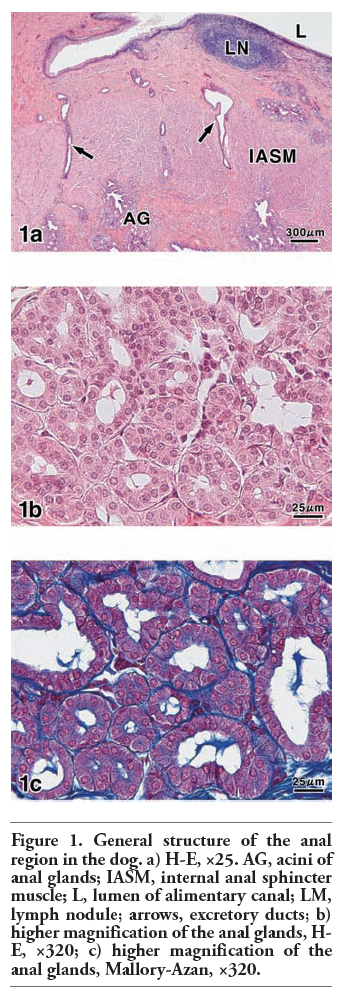 |
Figure 1. General structure of the anal region in the dog. a) H-E, ×25. AG, acini of anal glands; IASM, internal anal sphincter muscle; L, lumen of alimentary canal; LM, lymph nodule; arrows, excretory ducts; b) higher magnification of the anal glands, H-E, ×320; c) higher magnification of the anal glands, Mallory-Azan, ×320. |
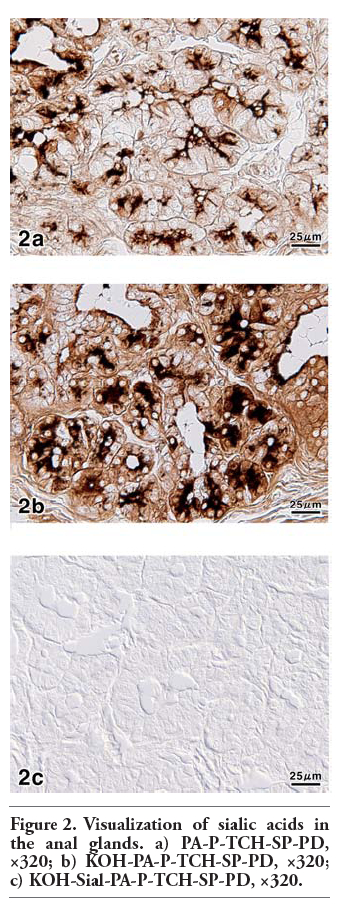 |
Figure 2. Visualization of sialic acids in the anal glands. a) PA-P-TCH-SP-PD, ×320; b) KOH-PA-P-TCH-SP-PD, ×320; c) KOH-Sial-PA-P-TCH-SP-PD, ×320. |
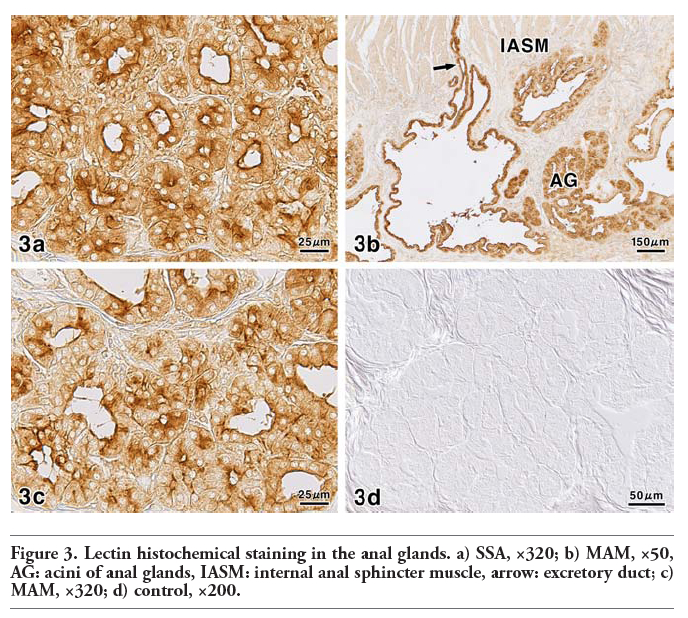 |
Figure 3. Lectin histochemical staining in the anal glands. a) SSA, ×320; b) MAM, ×50, AG: acini of anal glands, IASM: internal anal sphincter muscle, arrow: excretory duct; c) MAM, ×320; d) control, ×200. |
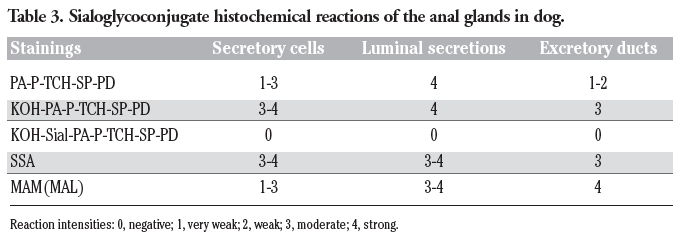 |
Table 3. Sialoglycoconjugate histochemical reactions of the anal glands in dog. |
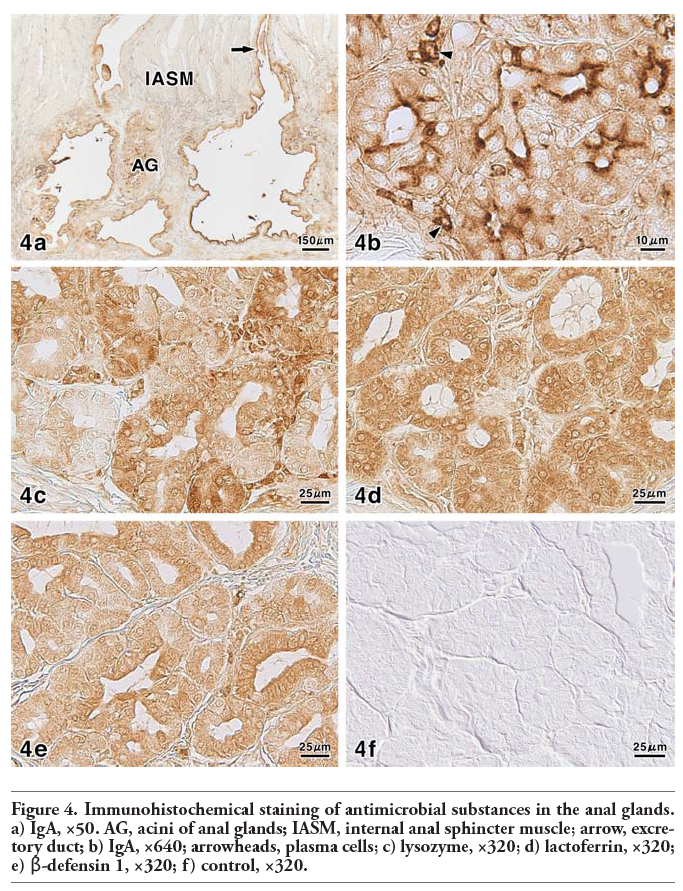 |
Figure 4. Immunohistochemical staining of antimicrobial substances in the anal glands. a) IgA, ×50. AG, acini of anal glands; IASM, internal anal sphincter muscle; arrow, excretory duct; b) IgA, ×640; arrowheads, plasma cells; c) lysozyme, ×320; d) lactoferrin, ×320; e) β-defensin 1, ×320; f) control, ×320. |
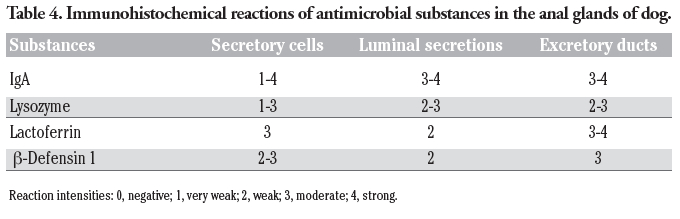 |
Table 4. Immunohistochemical reactions of antimicrobial substances in the anal glands of dog. |
In mammals, many studies have revealed that the salivary glandular secretions protect the oral cavity as the entrance of the digestive system.33,34,35 In contrast, there have been few studies with regard to the functional properties of the secretory products elaborated by the anal glands, which are distributed in the terminal region of the alimentary tract. According to our previous study, the secretory epithelium of the canine anal glands and their excretory duct system contain large amounts of neutral glycoproteins.8 In particular, α-L-fucose residues, which are prominent components of the glandular secretions, may modify or control the viscoelastic properties of the mucus in order to form a stable mucous coat of the rather dry feces. Additionally, the excretory ducts systems are deeply involved in the maturation of the secretions. The present examination disclosed the distribution of sialoglycoconjugates and antimicrobial substances in these glands, as supported by the PA-P-TCH-SP-PD, lectin histochemical and immunohistochemical procedures.
Saponification caused an increase of PA-P-TCH-SP-PD reactivity, indicating the presence of sialic acids with O-acetylated substitutions at C-8 and/or C-9.29,30,31 In addition, lectin histochemical results of SSA and MAM represent the localization of sialic acid residues, linked to α2-6Gal/GalNAc and α2-3Gal1-4GlcNAc, respectively. O-Acetylated sialic acids are known to reduce sialidase activity and extend the lifespan of serum glycoproteins.14,15 Sialoderivatives, with variation in their degree of acetylation and the types of linkages and acceptor sugars, may play an important role in defense against pathogenic agents,14,36,37 because, for example, influenza viruses differ in their ability to recognize Sia-Gal linkages depending on the animal hosts.38
The antimicrobial substances identified during the course of our study are generally part of the innate immunity system and of great importance for mucosal integrity by their antibacterial activities. They are widely represented in mucous secretions, including tears, saliva, colostrum and secretions from the genitourinary tract, gastrointestinal tract, prostate and respiratory epithelium. In this context, lysozyme is widely known as a remarkable bacteriolytic element found in tissues and secretions.39 This enzyme is capable of direct bacteriolytic action because it hydrolyzes β-(1,4)-glycosidic bonds between N-acetylnumamic acid and N-acetyl-β-D-glucosamine, which are commonly present in the peptidoglycan of bacterial cell walls.40,41 Furthermore, lysozyme has a close functional relationship to immunoglobulins and lactoferrin.41 Secretory IgA consists of polymeric IgA containing J chain and a distinctive polypeptide known as secretory component, and is a predominant immunoglobulin in external secretions of humans and various other mammals.42 IgA with J chain is produced by plasma cells. Then, secretory IgA is transported through the lateral borders and apically in the cytoplasm of secretory epithelial cells into external secretions.43,44,45 Our results are consistent with the above-mentioned view of the secretory mechanism of IgA. Lactoferrin has a broad spectrum of antimicrobial activity against bacteria, fungi and viruses.46,47 The antimicrobial properties of lactoferrin are imparted by two distinct mechanisms. The bacteriostatic function is caused by the strong iron binding properties of protein.48,49 On the other hand, lactoferrin also possesses bacteriocidal function because of a direct antimicrobial effect against microorganisms.50,51,52,53 In addition, lactoferrin inhibits allergen-induced skin inflammation, which is secondary to its role in regulating the production of cytokines.54,55 In mammals, β-defensins are produced by a variety of epithelial cells and phagocytes, and often present at high concentration.56,57 They have the capability of insertion into cell membranes, depending on their electrical forces. Consequently, most defensins cause the destruction of microorganisms, for example, bacteria, fungi and viruses.17,18,56,57 The canine β-defensin gene family consists of 43 members on the basis of sequence similarity with other mammalian β-defensins.48 The gene lineage and amino acid identity of canine β-defensin 1 resemble those of human β-defensin 1.58,59
In conclusion, the present study revealed the presence and distribution of sialoglycoconjugates and antimicrobial substances in the anal glands of dog. It is suggested from our findings that the glandular secretions elaborated by the canine anal glands are deeply involved not only in regulation of the mucous viscoelastic properties or scent marking but also in effective defense activities in the anal mucosa, as the terminal segment of the digestive tract. Such features are important to understand the biologically relevant characteristics of this gland type in mammalian species.
1. Ortmann R. Die Analregion der Säugetiere. Handb Zool 1960;8:1-68.
2. Gerisch D, Neurand K. Topographie und Histologie der Drüsen der Regio analis des Hundes. Zbl Vet Med C 1973;2:280-94.
3. Meyer W, Neurand K. Untersuchungen zur Histochemie der Glandulae anales des Hundes. Zbl Vet Med A 1981; 28:420-5.
4. Neurand K, Meyer W. Die Drüsen der Analregion des Hundes. Tierärztl Praxis 1982;10:243-52.
5. Budsberg SC, Spurgeon TL. Microscopic anatomy and enzyme histochemistry of the canine anal canal. Anat Histol Embryol 1983;12:295-316.
6. Schwarz R, Meyer W. Haut und Hautorgane. In: Frewein J, Vollmerhaus B (eds.) Die Anatomie von Hund und Katze. 1994, P. Parey Ed., Hamburg, Berlin, Germany, pp. 316-40.
7. Meyer W, Tsukise A, Neurand K. SEM and carbohydrate histochemical aspects of the glands in the anal region of the pig. Z Säugetierkunde 1983;48:245-55.
8. Tsukise A, Meyer W, Nagaoka D, Kikuchi K, Kimura J, Fujimori O. Lectin histochemistry of the canine anal glands. Ann Anat 2000;182:151-9.
9. McColl I. The comparative anatomy and pathology of anal glands. Ann R Coll Surg Engl 1967;40:36-67.
10. Fenger C, Filipe MI. Pathology of the anal glands with special reference to their mucin histochemistry. Acta Pathol Microbiol Scand A 1977;85:273-85.
11. Fenger C, Filipe MI. Mucin histochemistry of the anal canal epithelium. Studies of normal anal mucosa and mucosa adjacent to carcinoma. Histochem J 1981:13;921-30.
12. Fenger C. Histology of anal canal. Am J Surg Pathol 1988;12:41-55.
13. Klosterhalfen B, Offner F, Vogel P, Kirkpatrick CJ. Anatomic nature and surgical significance of anal sinus and anal intramuscular glands. Dis Colon Rectum 1991:34;156-60.
14. Schauer R. Sialic acids as regulators of molecular and cellular interactions. Curr Opin Struct Biol 2009;19:507-14.
15. Varki A, Schauer R. Sialic acids. In: Varki A, Cummings RD, Esko JD, Freeze HH, Stanley P, Bertozzi CR, Hart GW, Etzler ME (eds.) Essentials of Glycobiology. 2nd ed, 2009 Cold Spring Harber Laboratory Press, New York, USA, pp. 199-218.
16. Schröder JM. Epithelial peptide antibiotics. Biochem Pharmacol 1999;57:121-34.
17. Bos JD, Pasch MC, Asghar SS. Defensins and complement systems from the perspective of skin immunity and autoimmunity. Clin Dermatol 2001;19:563-72.
18. Yang D, Chertov O, Oppenheim JJ. The role of mammalian antimicrobial peptides and proteins in awakening of innate host defense and adaptive immunity. Cell Mol Life Sci 2001;58:978-89.
19. Meyer W, Neurand K, Tanyola A. General anti-microbial properties of the integument in fleece producing sheep and goats. Small Rumin Res 2001;41:181-90.
20. Yasui T, Tsukise A, Fukui K, Kuwahara K, Meyer W. Aspects of glycoconjugate production and lysozyme- and defensin-expression of the ceruminous glands of the horse (Equus prezewalskii f. dom.). Eur J Morphol 2005;42:127-34.
21. Yasui T, Fukui K, Nara T, Habata I, Meyer W, Tsukise A. Immunocytochemical localization of lysozyme and beta-defensin in the apocrine glands of the equine scrotum. Arch Dermatol Res 2007;299:393-7.
22. Yasui T, Nara T, Habata I, Meyer W, Tsukise A. Functional properties of feline foot pads as studied by lectin histochemical and immunohistochemical methods. Acta Histochem 2010;112:169-77.
23. Ueda T, Fujimori O, Yamada K. A new histochemical method for detection of sialic acids using a physical development procedure. J Histochem Cytochem 1995;43: 1045-51.
24. Spicer SS. A correlative study of the histochemical properties of rodent acid mucopolysaccharides. J Histochem Cytochem 1960;8:18-35.
25. Reid PE, Park CM. Carbohydrate histochemistry of epithelial glycoproteins. Prog Histochem Cytochem 1990;21:1-170.
26. Spicer SS, Horn RG, Leppi TJ. Histochemistry of connective tissue mucopolysaccharides. In: Wagner BM, Smith DE (eds.) The connective tissue. 1967, Williams and Wilkens, Baltimore, USA, pp. 251-303.
27. Uchida Y, Tsukada Y, Sugimori T. Enzymatic properties of neuraminidase from Arthrobacter urefaciens. J Biochem 1979;86:1573-5.
28. Schauer R. Achevements and challenges of sialic acid research. Glycoconj J 2000;17: 485-99.
29. Ueda T, Fujimori O, Tsukise A, Yamada K. Histochemical analysis of sialic acids in the epididymis of the rat. Histochem Cell Biol 1998;109:399-407.
30. Yasui T, Tsukise A, Fukui K, Kuwahara Y, Meyer W. Histochemical localization of complex carbohydrates in the nasolabial glands of the Japanese deer (Cervus nippon yakushimae). Acta Histochem: 2005; 107, 187-198.
31. Habata I, Yasui T, Tsukise A. Histochemistry of sialoglycoconjugates in goat submandibular glands. Anat Histol Embryol 2011 40:187-195.
32. Danguy A. Perspectives in modern glycohistochemistry. Eur J Bas Appl Histochem 1995;39:5-14.
33. Nieuw Amerongen AV, Bolscher JG, Veerman EC. Salivary proteins: protective and diagnostic value in cariology? Caries Res 2004;38:247-53.
34. Scocco P, Pedini V. Equine mandibular gland: in situ characterisation of sialoderivatives. Equine Vet J 2006;38:410-5.
35. Tabak LA. In defense of the oral cavity: the protective role of the salivary secretions. Pediatr Dent 2006:28;110-7.
36. Schauer R. Sialic acids: fascinating sugars in higher animals and man. Zoology 2004;107:49-64.
37. Parillo F, Arias MP, Supplizi AV. Glycoprofile of the different cell types present in the mucosa of the horse guttural pouches. Tissue Cell 2009;41:257-65.
38. Suzuki Y. Sialobiology of influenza molecular mechanism of host range variation of influenza viruses. Biol Pharm Bull 2005; 28:399-408.
39. Fleming A. On a remarkable bacteriolytic element found in tissues and secretions. Proc R Soc Lond B Biol Sci 1922;93:306-10.
40. Strominger JL, Tipper DJ. Structure of bacterial cell walls: the lysozyme substrate. In: Osserman EF, Canfield RE, Beychok S (eds.) Lysozyme. 1974, Academic Press, New York, USA, pp. 169-85.
41. Jolles P, Jolles J. What’s new in lysozyme research? Always a model system, today as yesterday. Mol Cell Biochem 1984;63:165-89.
42. Tang QJ, Tao KZ, Yun-Liu, Sun XJ, Geng MY, Jiang CL. Immunocytochemical localization of secretory component in Paneth cell secretory granules-rat Paneth cells participate in acquired immunity. J Mol Histol 2005;36:331-5.
43. Brandtzaeg P. Mucosal and glandular distribution of immunoglobulin components: differential localization of free and bound SC in secretory epithelial cells. J Immunol 1974;112:1553-9.
44. Brandtzaeg P. Immunohistochemical studies on various aspects of glandular immunoglobulin transport in man. Histochem J 1977;9:553-72.
45. Kaetzel CS, Robinson JK, Chintalacharuvu KR, Vaerman JP, Lamm ME. The polymeric immunoglobulin receptor (secretory component) mediates transport of immune complexes across epithelial cells: a local defense function for IgA. Proc Natl Acad Sci USA 1991;88:8796-800.
46. Levay PF, Viljoen M. Lactoferrin: a general review. Haematologica 1995;80:252-67.
47. Valenti P, Marchetti M, Superti F, Amendolia MG, Puddu P, Gessani S, et al. Antiviral activity of lactoferrin. Adv Exp Med Biol 1998;443:199-203.
48. Bullen JJ, Rogers HJ, Griffiths E. Role of iron in bacterial infection. Curr Top Microbiol Immunol 1978;80:1-35.
49. Bezwoda WR, Mansoor N. Lactoferrin from human breast milk and from neutrophil granulocytes. Comparative studies of isolation, quantitation, characterization and iron binding properties. Biomed Chromatogr 1989;3:121-6.
50. Arnold RR, Cole MF, McGhee JR. A bacterial bactericidal effect for human lactoferrin. Science 1977;197:263-5.
51. Bellamy W, Takase M, Wakabayashi H, Kawase K, Tomita M. Antibacterial spectrum of lactoferricin B, a potent bactericidal peptide derived from the N-terminal region of bovine lactoferrin. J Appl Bacteriol 1992;73:472-9.
52. Bellamy W, Takase M, Yamauchi K, Wakabayashi H, Kawase K, Tomita M. Identification of the bactericidal domain of lactoferrin. Biochim Biophys Acta 1992;1121:130-6.
53. Tomita M, Takase M, Bellamy W, Shimamura S. A review: the active peptide of lactoferrin. Acta Paediatr Jpn 1994;36:585-91.
54. Kimber I, Cumberbatch M, Dearman RJ, Headon DR, Bhushan M, Griffiths CE. Lactoferrin: influences on Langerhans cells, epidermal cytokines, and cutaneous inflammation. Biochem Cell Biol 2002;80: 103-7.
55. Ward PP, Uribe-Luna S, Conneely OM. Lactoferrin and host defense. Biochem Cell Biol 2002;80:95-102.
56. Ganz T. Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol 2003;3:710-20.
57. Ganz T. Defensins: antimicrobial peptides of vertebrates. C R Biol 2004;327:539-49.
58. Patil AA, Chai Y, Sang Y, Blecha F, Zhang G. Cross-species analysis of the mammalian beta-defensin gene family: presence of syntenic gene clusters and preferential expression in the male reproductive tract. Physiol Genomics 2005;23:5-17.
59. Wingate KV, Torres SM, Silverstein KA, Hendrickson JA, Rutherford MS. Expression of endogenous antimicrobial peptides in normal canine skin. Vet Dermatol 2009;20:19-26.
[TOP]