
K. Fukui,1 T. Yasui,1 H. Gomi,1 H. Sugiya,2 O. Fujimori,3 W. Meyer,4 A. Tsukise1
1Department of Veterinary Anatomy, College of Bioresource Sciences, Nihon University, Fujisawa, Kanagawa, Japan; 2Laboratory of Veterinary Biochemistry, College of Bioresource Sciences, Nihon University, Kanagawa, Japan;
3Laboratory of Anatomy and Histochemistry, Faculty of Rehabilitation Science, Nagoya Gakuin University, Seto, Aichi, Japan; 4Institute for Anatomy, University of Veterinary Medicine Hannover Foundation, Hannover, Germany
Correspondence: Prof. Azuma Tsukise, Department of Veterinary Anatomy, College of Bioresource Sciences, Nihon University, 1866 Kameino, Fujisawa, Kanagawa 252-0880, Japan. Tel. +81.466.843629 - Fax: +81.466.843629.
E-mail: tsukise@brs.nihon-u.ac.jp
Key words: sialic acids, antimicrobial substances, eccrine glands, porcine snout skin, histochemistry.
Received for publication: 5 October 2011.
Accepted for publication: 15 December 2011.
©Copyright K. Fukui et al., 2012
Licensee PAGEPress, Italy
European Journal of Histochemistry 2012; 56:e6
doi:10.4081/ejh.2012.e6
AbstractThe distribution of sialic acids and antimicrobial products (lysozyme, IgA, lactoferrin, β-defensin 2) as well as Rab3D in the eccrine glands of porcine snout skin was studied by sialoglycoconjugate histochemistry and immunohistochemistry. The secretory epithelium consisted of two types of secretory cells: dark and clear cells. The dark cells exhibited considerable amounts of sialoglycoconjugates, which included O-acetylated sialic acids, whereas sialic acids in the sequence Siaα2-3Gal1-4GlcNAc were confined to some of the dark cells. All antimicrobial substances and Rab3D were demonstrated to be also mainly present in some of the dark cells. Additionally, in the cytological and cytochemical features, the different characteristics were observed among the dark cells. The results obtained are discussed with regard to the functional significance of the eccrine glands. The secretory products elaborated by this gland type may function as protective agents in order to preserve the skin integrity of the snout region, considering that sialic acids and antimicrobial substances are important in general defense mechanisms. |
In most mammals, except for humanoid primates, apocrine glands are extensively distributed throughout the common integument, whereas eccrine glands are confined to the skin of a series of specific body regions, for example, the foot pads of carnivores, the frog of ungulates (Cuneus ungulae), the carpus of pig and the nasolabial region of ruminants and pig.1,2,3,4,5,6,7 These specific regions have been modified by structural and functional adaptations that enable the animal to meet the biological requirements of its environment. The snout skin of the pig, for example, one of such structures that have undergone modifications, can function as a delicate sense organ. The eccrine glands of this region were found to contain considerable amounts of neutral glycoproteins, especially in the dark cells. It is considered that these products are closely related to the specific integumental functions.8
It is widely known that sialic acids are mainly located at the terminal position of glycoproteins. They have different functional properties depending on the degree of acetylation and on the linkage of sialic acid to the acceptor sugars. Sialoderivatives seem to perform a key role as the most versatile function modulators in cell biology and pathology.9,10 The skin is a first barrier against pathogens. In this context, in several mammalian groups, recent studies have revealed that different antimicrobial substances are distributed also in eccrine glands.11-13 Furthermore, the immune system has developed a special integumental subsystem to bind all of the possible microbial invaders and their products, to present them and to eliminate them rapidly.14,15,16
The present study on the eccrine glands of porcine snout skin investigates the localization of sialic acids and antimicrobial substances using lectin histochemical and immunohistochemical methods. Thus, our findings may be important for understanding the normal functions of snout skin in the pig independent of its abilities as a sensory organ.
All experiments were performed in accordance with the guidelines for the care and use of laboratory animals at the Institute of Experimental Animal Science, College of Bioresource Sciences, Nihon University. Five male miniature pigs (potbelly, 1-2 years, 40-50 kg) were deeply anesthetized and then exsanguinated from the common carotid arteries. After bloodletting, the snout skin was removed surgically. These skin samples were fixed in Bouin’s solution for 24 h at room temperature, then carefully washed and dehydrated through a graded series of ethanol. The materials were embedded in paraffin wax and then sectioned at a thickness of 5 mm. Sections were stained with haematoxylin-eosin (H-E), Mallory-Azan and alcian blue (AB) pH 2.5-periodic acid Schiff (PAS)17 methods for general histological purposes, as well as with sialoglycoconjugate histochemical and immunohistochemical procedures.
For the localization of sialic acid residues, after deparaffinization, the histochemical staining method employed was AB pH 2.518 and periodic acid-phenylhydrazinium chloride-thiocarbohydrazide-silver proteinate-physical development (PA-P-TCH-SP-PD).19 Selective periodate oxidation (0.4 mM PA in 1 M HCl) of sialic acid residues yields monoaldehyde at C-7 or C-8 because it oxidizes vicinal diols at C-7 and C-8 or at C-8 and C-9. Then, monoaldehyde condenses with phenylhydrazine to give rise to the corresponding phenylhydrazone. This compound can undergo solvolysis to yield TCH-SP-PD-reactive monoaldehydes. On the other hand, the dialdehydes of neutral sugars oxidized by selective periodate oxidation condense with phenylhydrazine to result in TCH-SP-PD-unreactive azido or morpholine. In addition, sialic acids with O-acetyl substitutions at C-8 and/or C-9 are undetected by this procedure. Therefore, chemical modification by saponification (KOH) (0.5% potassium hydroxide-70% ethanol)20 was performed on some sections, prior to PA-P-TCH-SP-PD staining, since KOH treatment is known to remove the acetyl groups on C-4, C-7, C-8 and C-9. Enzyme digestion with sialidase (from Arthrobacter ureafaciens, Nacalai Tesque Inc., Kyoto, Japan; 1.0 U/mL in 0.1 M phosphate buffer, pH 5.3, at 37˚C for 18 h)21 after saponification was conducted before the AB pH 2.5 and PA-P-TCH-SP-PD stainings. These physical development procedures for detection of sialic acids and their moieties are summarized in Table 1.
Additionally, the sections were incubated with biotinylated lectins at concentrations of 10-20 mg/mL in 0.05 M phosphate-buffered saline (PBS) (pH 7.2) for 60 min at room temperature, following treatment with 0.3% hydrogen peroxide to suppress endogenous peroxidase (PO) activity and then preincubation with 1% bovine serum albumin (BSA) in PBS for 30 min at room temperature. The lectins used were wheat germ agglutinin (WGA), Sambucus sieboldiana agglutinin (SSA) and Maackia amurensis agglutinin (MAM) (Seikagaku Kogyo Co., Tokyo, Japan). Their specificities and inhibitory sugars are listed in Table 2.22 After rinsing with PBS, the sections were soaked in PO-labeled streptavidin (Nichirei Co., Tokyo, Japan) for 30 min at room temperature. The activity of the PO employed for labeling was evaluated with a solution containing 0.05 M Tris-HCl buffer (pH 7.6), diaminobenzidine (DAB) and hydrogen peroxide. Lectin controls were prepared by addition of inhibitory sugars at a final concentration of 0.01 M to the respective lectin solutions, by substitution of unconjugated lectins for biotinylated lectin conjugates or by exposure of sections to the PO-DAB system without lectin.
For the immunohistochemical methods, the sections were pretreated with 0.3% hydrogen peroxide for 30 min for the blocking of endogenous PO activities. Then, they were incubated with primary antibodies diluted in 0.01 M PBS (pH 7.3) against lysozyme (polyclonal, from rabbit) (Dako, Glostrup, Denmark), IgA (polyclonal, from rabbit) (Dako), lactoferrin (polyclonal, from rabbit) (MP Biomedicals, Ohio, USA), human β-defensin 2 (polyclonal, from rabbit) (Biolog, Kronshagen, Germany) and Rab3D (polyclonal, from rabbit) (Proteintech, Illinois, USA) for 24 h at 4°C, following preincubation with 1% BSA in PBS for 30 min at room temperature. The primary antibodies were diluted as follows: lysozyme 1:100, IgA 1:100, lactoferrin 1:100, human β-defensin 2 1:400, Rab3D 1:50. For antigen retrieval, some of the sections were incubated with Proteinase K solution (Dako) for 3 min at room temperature before incubation with the primary antibodies to lysozyme, IgA, lactoferrin and β-defensin 2. With regard to anti-Rab3D, the sections were treated with microwave irradiation (500 W, for 6 min) in 0.01 M citrate buffer at pH 6.0. After rinsing with PBS, these sections were incubated with prediluted dextran-polymer-conjugated secondary antibodies (EnVision+ Dual Link System-HRP, raised in goat against mouse and rabbit immunoglobulins, Dako) for 30 min at room temperature. The immunoreaction employed for labeling was revealed using a PO-DAB system, as described above. Controls for immunohistochemical methods were performed by incubation with PBS without primary antibodies or exposure of sections to the PO-DAB system without primary or secondary antibodies.
For general cytology, tissue pieces were fixed in 2.5% glutaraldehyde (GA) solution in 0.1 M phosphate-buffered solution (PB) (pH 7.4) for 2 h at 4°C, post-fixed in 2% osmium tetroxide solution and embedded in Epon 812.23 From these tissue blocks, ultrathin sections were cut on an ultramicrotome, mounted on copper grids and stained with uranyl acetate24 and lead citrate.25 For the cytochemical demonstration, the tissue specimens were fixed in a mixture of 4% paraformaldehyde (PFA) and 0.5% GA in 0.1 M PB (pH 7.4) for 2 h at 4˚C, and embedded in LR-White resin.26 For LR-White-embedded materials, ultrathin sections were cut as stated above and placed on nylon grids. The sections on nylon grids were subjected to the periodic acid-thiocarbohydrazide-silver proteinate-physical development procedure (PA-TCH-SP-PD), which is an efficient and sensitive cytochemical method for the detection of vicinal diols of neutral carbohydrates.27 These sections were then counterstained with uranyl acetate and lead citrate. For the cytochemical identification of glycogen in the cytoplasm, enzyme digestion with α-amylase (from Bacillus subtilis, Seikagaku Kogyo Co., 1 mg/mL in 0.1 M PBS pH 6.0, at 37°C for 4 h)28 was carried out on some sections prior to the PA-TCH-SP-PD procedure.
In pig snout skin, the eccrine glands were visualized in the connective tissue of the hypodermis. Their secretory epithelium was commonly distinguished from dark cells and clear cells (Figure 1 a,b). In the sections stained with AB pH 2.5-PAS, however, different reaction intensities were clearly detected among the dark cells, which showed moderate or strong reactions (Figure 2 a,b). The excretory duct penetrated toward the epidermis and directly opened onto the skin surface.
In the eccrine glandular acini, the AB pH 2.5 procedure gave rise to a weak to moderate reaction of the dark cells and a very weak reaction of the clear cells (Figure 3a). Enzyme digestion with sialidase following saponification caused a weaker AB pH 2.5 reaction (Figure 3b). The PA-P-TCH-SP-PD procedure produced a weak to moderate reaction in the dark cells and a very weak reaction in the clear cells (Figure 3c). Saponification clearly increased the intensity of the PA-P-TCH-SP-PD reaction, in that the reaction intensity of the dark cells was markedly altered (Figure 3 d,e). The PA-P-TCH-SP-PD staining was nearly negative after sialidase digestion following saponification (Figure 3f). As for lectin histochemical methods, the WGA and SSA techniques resulted in a moderate to strong reaction of the dark cells, and a very weak reaction of the clear cells (Figure 4 a,b). The luminal secretions exhibited moderate reactions after the application with WGA and SSA. When incubated with MAM, a distinct reaction was detected in the luminal surface of some of the dark cells. Additionally, the cytoplasm of this cell type showed a weak reaction. On the other hand, the other dark cells and clear cells were almost negative (Figure 4c). In all of the control experiments for the lectin histochemical procedure, positive reactions were greatly diminished or nearly abolished in the eccrine glands (Figure 4d). The results obtained from the sialoglycoconjugate histochemical procedures are summarized in Table 3.
With regard to the immunohistochemical approach for lysozyme, some of the dark cells were found to exhibit a moderate to strong positive reaction, while the other dark cells and clear cells showed very weak reactions (Figure 5a). A weak to moderate positive reaction was observed in the luminal secretions with antibodies to lysozyme. Antibodies to IgA produced a very weak to weak positive reactions in some of the dark cells and in the luminal secretions, whereas the secretory epithelial cells except for these dark cells were almost negative (Figure 5b). In addition, a positive reaction was clearly detectable in the lumina of the blood vessels surrounding the secretory portion of the eccrine glands. Incubation with antibodies to lactoferrin and β-defensin 2 led to the same staining images as those obtained with antibodies to lysozyme in that the reaction intensity of some of the dark cells showed distinct positive reactions (Figure 5 c,d). Antibodies to Rab3D gave rise to a weak to moderate reaction of some of the dark cells, while the clear cells and luminal secretions were seen to react very weakly (Figure 5e).
In control experiments for the immunohistochemical methods performed by incubation with PBS without primary antibodies or exposure of sections to the PO-DAB system without primary or secondary antibodies, no glandular structures exhibited any positive reactions (Figure 5f). These results are summarized in Table 4.
Transmission electron microscopy of the eccrine glands of porcine snout skin confirmed that the secretory portion consisted of dark cells and clear cells with associated with myoepithelial cells. The secretory cells rested on a basement membrane and their nuclei were located in the central or basal region of the cytoplasm. Additionally, different morphological characteristics were detected among the dark cells. Therefore, we expediently classified the respective dark cells as type I dark cells and type II dark cells (Figure 6 a,b). Both cell types of the dark cells contained a well-developed Golgi apparatus and rough-surfaced endoplasmic reticulum within their cytoplasm. Although varying numbers of secretory granules possessing medium electron density were visible in the apical cytoplasm of these dark cells, the secretory granules of the type II dark cells were smaller than those of the type I dark cells (Figure 6b). On the other hand, the clear cells, the cytoplasm of which exhibited a lower electron density, were equipped with a Golgi apparatus, and contained large amounts of glycogen particles distributed throughout the cytoplasm. The cytoplasmic matrix of these secretory epithelial cells, especially of the clear cells, was studded with many mitochondria of varying morphology.
In the secretory cells stained with the PA-TCH-SP-PD procedure, the surface coat of the plasma membrane and the basement membrane exhibited positive reactions (Figure 7a-c). The prominent PA-TCH-SP-PD-reactive structures in the dark cells were the secretory granules and cisternae of the Golgi apparatus. However, in the dark cells, the secretory granules of the type I cells showed a weak to moderate reaction, whereas a distinct positive reaction was observed in those of the type II cells (Figure 7a, c). The PA-TCH-SP-PD-reactive ultrastructures in the clear cells were glycogen particles and also cisternae of the Golgi apparatus (Figure 7 a,b). Digestion with α-amylase abolished the PA-TCH-SP-PD-reactive glycogen particles, found particularly in the cytoplasm of the clear cells.
|
Table 1. The moieties detected by sialoglycoconjugate histochemical procedures. |
 |
Table 2. The lectins used and their receptor sugars and inhibitory sugars. |
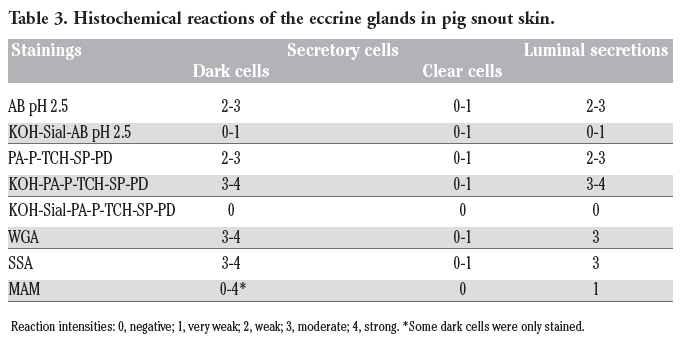 |
Table 1. The Table 3. Histochemical reactions of the eccrine glands in pig snout skin. detected by sialoglycoconjugate histochemical procedures. |
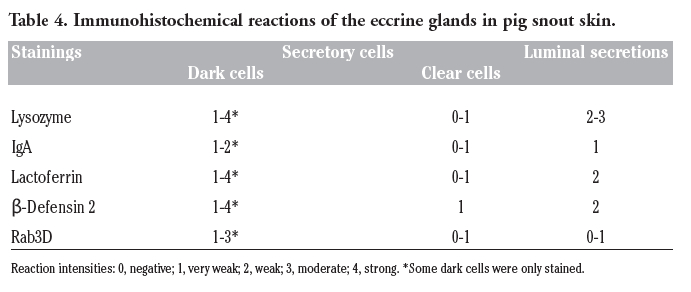 |
Table 4. Immunohistochemical reactions of the eccrine glands in pig snout skin. |
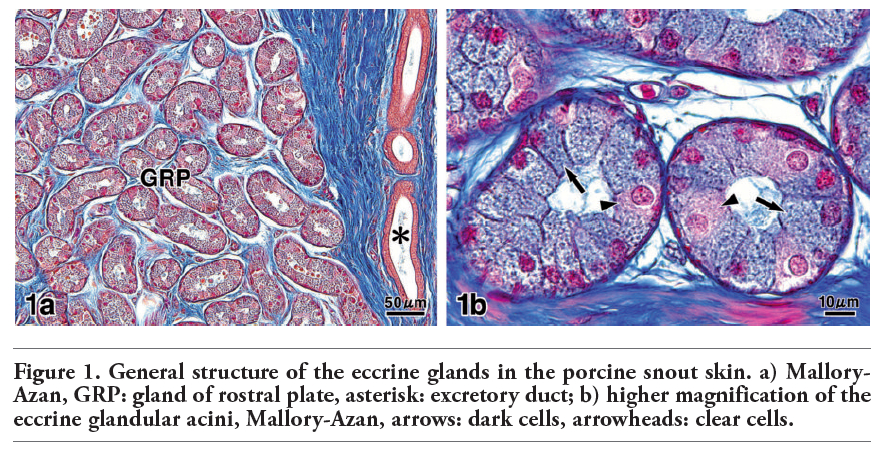 |
Figure 1. General structure of the eccrine glands in the porcine snout skin. a) Mallory-Azan, GRP: gland of rostral plate, asterisk: excretory duct; b) higher magnification of the eccrine glandular acini, Mallory-Azan, arrows: dark cells, arrowheads: clear cells. |
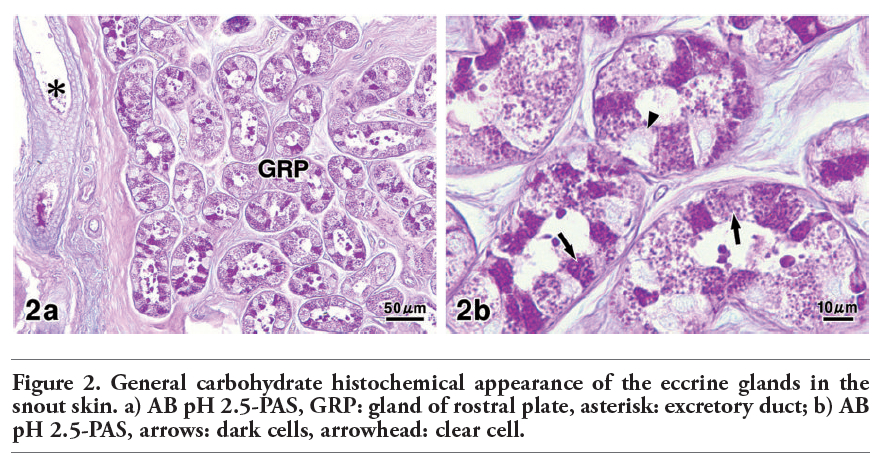 |
Figure 2. General carbohydrate histochemical appearance of the eccrine glands in the snout skin. a) AB pH 2.5-PAS, GRP: gland of rostral plate, asterisk: excretory duct; b) AB pH 2.5-PAS, arrows: dark cells, arrowhead: clear cell. |
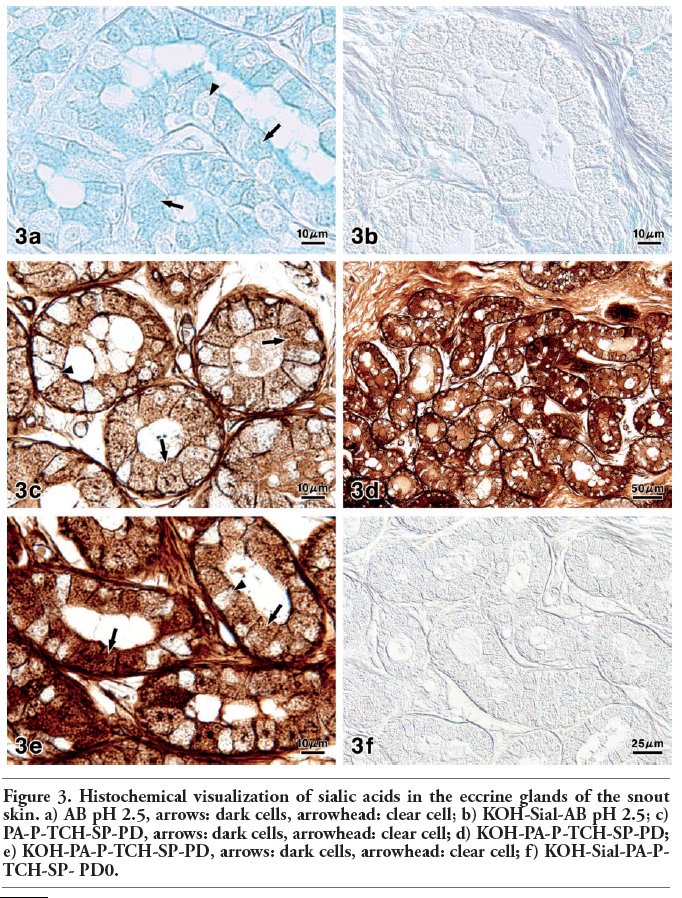 |
Figure 3. Histochemical visualization of sialic acids in the eccrine glands of the snout skin. a) AB pH 2.5, arrows: dark cells, arrowhead: clear cell; b) KOH-Sial-AB pH 2.5; c) PA-P-TCH-SP-PD, arrows: dark cells, arrowhead: clear cell; d) KOH-PA-P-TCH-SP-PD; e) KOH-PA-P-TCH-SP-PD, arrows: dark cells, arrowhead: clear cell; f) KOH-Sial-PA-P-TCH-SP- PD0. |
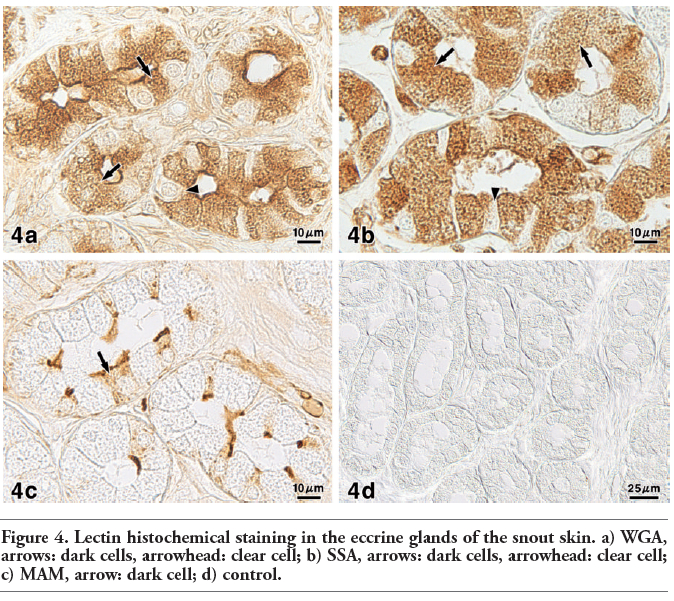 |
Figure 4. Lectin histochemical staining in the eccrine glands of the snout skin. a) WGA, arrows: dark cells, arrowhead: clear cell; b) SSA, arrows: dark cells, arrowhead: clear cell; c) MAM, arrow: dark cell; d) control. |
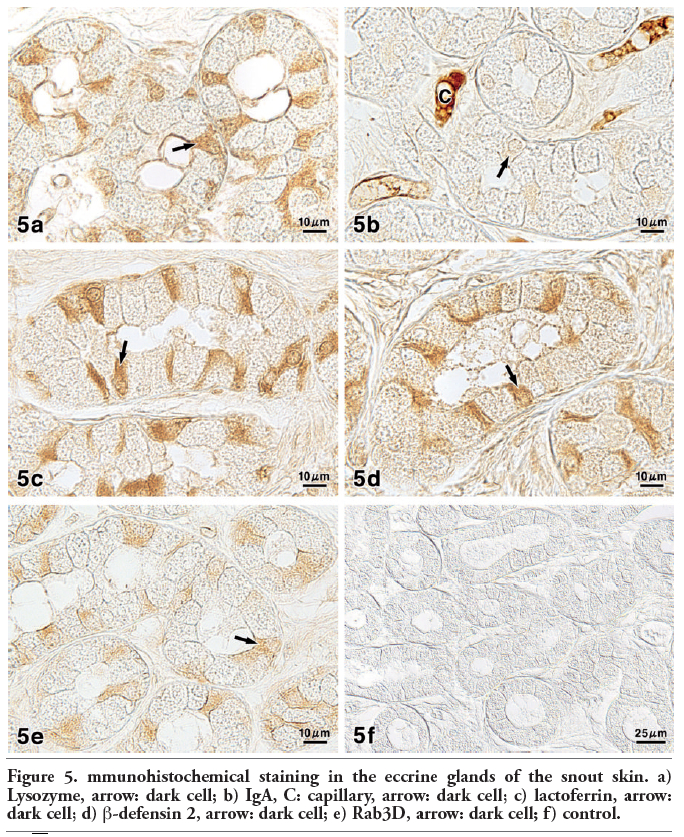 |
Figure 5. mmunohistochemical staining in the eccrine glands of the snout skin. a) Lysozyme, arrow: dark cell; b) IgA, C: capillary, arrow: dark cell; c) lactoferrin, arrow: dark cell; d) β-defensin 2, arrow: dark cell; e) Rab3D, arrow: dark cell; f) control. |
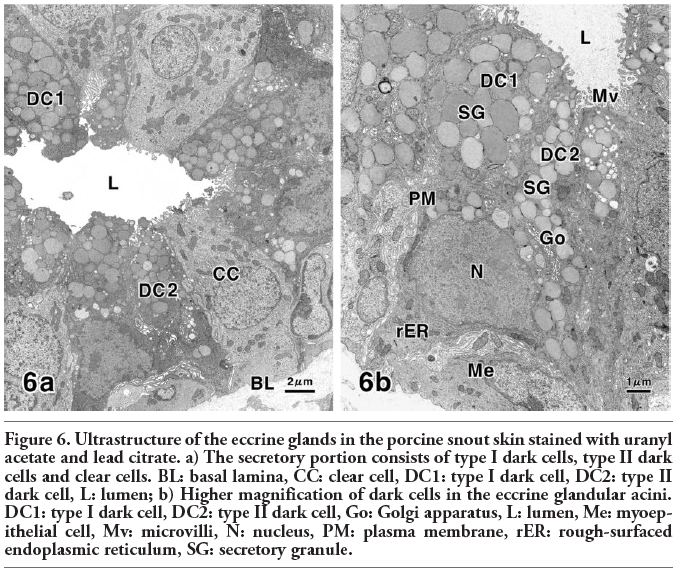 |
Figure 6. Ultrastructure of the eccrine glands in the porcine snout skin stained with uranyl acetate and lead citrate. a) The secretory portion consists of type I dark cells, type II dark cells and clear cells. BL: basal lamina, CC: clear cell, DC1: type I dark cell, DC2: type II dark cell, L: lumen; b) Higher magnification of dark cells in the eccrine glandular acini. DC1: type I dark cell, DC2: type II dark cell, Go: Golgi apparatus, L: lumen, Me: myoepithelial cell, Mv: microvilli, N: nucleus, PM: plasma membrane, rER: rough-surfaced endoplasmic reticulum, SG: secretory granule. |
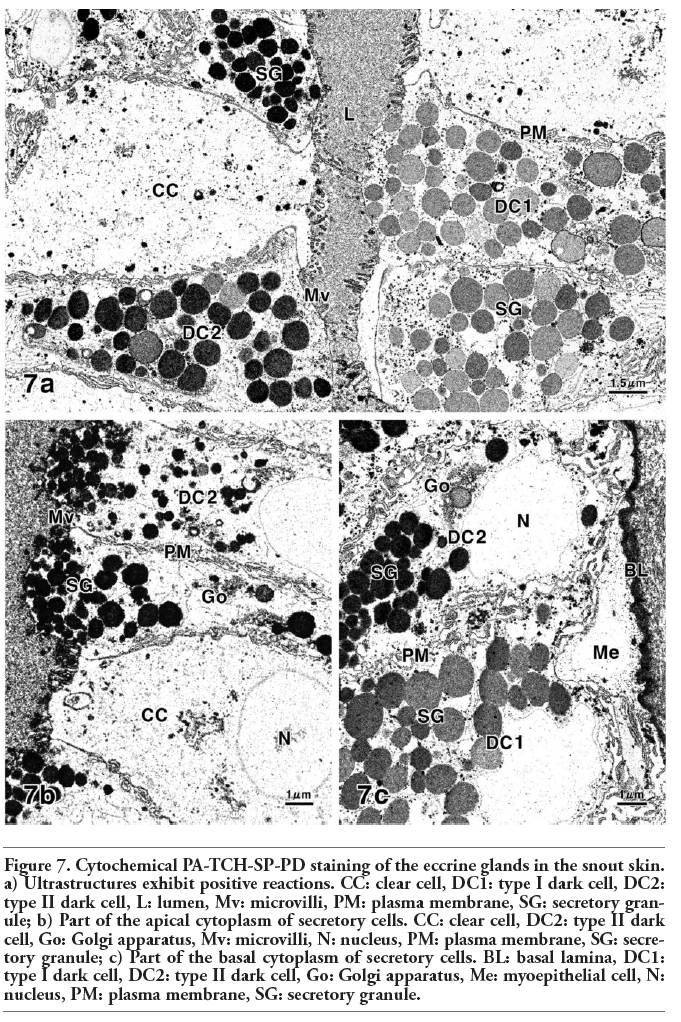 |
Figure 7. Cytochemical PA-TCH-SP-PD staining of the eccrine glands in the snout skin. a) Ultrastructures exhibit positive reactions. CC: clear cell, DC1: type I dark cell, DC2: type II dark cell, L: lumen, Mv: microvilli, PM: plasma membrane, SG: secretory granule; b) Part of the apical cytoplasm of secretory cells. CC: clear cell, DC2: type II dark cell, Go: Golgi apparatus, Mv: microvilli, N: nucleus, PM: plasma membrane, SG: secretory granule; c) Part of the basal cytoplasm of secretory cells. BL: basal lamina, DC1: type I dark cell, DC2: type II dark cell, Go: Golgi apparatus, Me: myoepithelial cell, N: nucleus, PM: plasma membrane, SG: secretory granule. |
In various mammalian species including humans, the secretory cells of the eccrine glands are generally divided into dark cells and clear cells, which participate in secreting glycoproteins and ion fluid, respectively.5,11,12,29,30,31 Unlike in other mammalian species, however, the dark cells of the eccrine glands in the porcine snout skin could be classified morphologically and cytochemically as type I cells and type II cells, owing to the ultrastructural features and the results of the PA-TCH-SP-PD procedure. However, it is unclear whether their morphological and morphochemical heterogeneity depends on the different functional stages or maturation stages. Furthermore, our results revealed the presence of sialic acids and various antimicrobial substances in these glands.
Within the saccharide residues, the functional properties of sialic acids appear to be manifold.9,10 These glycoconjugates protect cells and macromolecules against enzymatic and immunological attacks by inhibiting the adherence of different bacteria, and can function as recognition sites for various receptors.9,10,32,33 These functions are also based on the presence of sialic acids with O-acetyl substitution at C-8 and/or C-9, as indicated by the increase of PA-P-TCH-SP-PD reactivity following saponification.34 Moreover, apoptosis was reported to be inhibited by O-acetylated sialic acids.35 In this study, the dark cells contained sialoglycoconjugates that terminated in Siaα2-6Gal/GalNAc, whereas sialic acids linked to α2-3Gal1-4GlcNAc were only localized in the luminal surface of some of the dark cells. Such distributional patterns of sialoglycoconjugates are clearly different from those in the eccrine glands of other mammals.7,13,30,31,36 The great variety of sialic acid residues, in particular, may play an important role in the general defense against pathogenic agents.10,37,38
Our findings showed that antimicrobial materials are also produced in the eccrine glands of porcine snout skin and released as part of the secretions. The localization of these substances was mainly confined to some of the dark cells in the eccrine glands. In humans, IgA, lactoferrin and β-defensins were immunohistochemically detected in the eccrine glandular cells.39,40,41 Although Ezoe and Katsumura42 reported that lysozyme was not present in human eccrine glands, another investigation identified the expression of this enzyme.43 With regard to this gland type of the feline foot pads, these antimicrobial substances were demonstrated to be immunolocalized in the dark cells.13 Moreover, in hyrax and tenrec, lysozyme and β-defensins are also observed in the eccrine glands of foot pads.11,12
Lysozyme is a 14.5-KD cationic enzyme with bactericidal activity that catalyzes the hydrolysis of glycosidic bonds in bacterial cell walls.44 This enzyme has a close functional relationship to immunoglobulins and lactoferrin.45 The predominant immunoglobulin isotype on most mucosal surfaces and external secretions is secretory IgA composed of polymeric IgA containing J chain and secretory component.46,47 In epithelial defense, IgA contributes to preventing pathogen adhesion to host cells, thereby blocking dissemination and further infection.48 IgA as well as J chain is produced by plasma cells and transported through secretory epithelial cells into external secretions.49 However, in the present study, the plasma cells were scarcely observed in the connective tissue surrounding the eccrine glandular acini. Furthermore, lactoferrin is a multifunctional glycoprotein of the transferrin family that is widely represented in various secretory fluids. It is one of the components of innate immunity because of its bacteriocidal, fungicidal and antiviral activities.50,51
Additionally, lactoferrin inhibits allergen-induced skin inflammation, which is secondary to its role in regulating the production of cytokines.52 In humans and other mammals, defensins are known as some of the main antimicrobial peptides, which attack the cell walls of a wide range of bacterial, fungal and viral pathogens by insertion in the phospholipid bilayer, leading to disruption and subsequent death of the microorganisms.53,54 This is most important for the integument regarding the multitude of microbes always active on the skin surface.55,56
It is established that Rab proteins function in the tethering or docking of vesicles to their target compartment, leading to membrane fusion.57 Rab3D is expressed in secretory granules of exocrine secretory cells, and involved in regulated exocytosis.58,59
The present study revealed that the secretory granules of the type II dark cells had a higher concentration of glycoproteins than those of the type I dark cells, as supported by the staining ability of the PA-TCH-SP-PD procedure.27 It is suggested from these findings that sialic acid residues linked to Siaα2-3Gal1-4GlcNAc and various antimicrobial substances are elaborated by the type II dark cells. Furthermore, the localization of Rab3D seems to be consistent with that of these products. Such features indicate that this regulatory protein plays an important role in the secretory regulation of them.
Summarizing our functional interpretations, the results of this study confirmed the ultrastructures and the distribution of sialoglycoconjugates, antimicrobial substances and Rab3D in porcine snout eccrine glands. Sialoglycoconjugates are involved not only in adsorption of pathogenic agents, but also in viscoelasticity of secretions.35 Moreover, the antimicrobial substances are important in the innate immune response.
In conclusion, the eccrine glandular secretions containing these secretory components may be essential for preserving the integrity of porcine snout skin as a sensory organ.
1. Ellis RA. Eccrine sweat glands: electron microscopy, cytochemistry and anatomy. In: J. Jadassohn (ed.) Handbuch der Haut- und Geschlechtskrankheiten, Ergänzungswerk I/1. 1968, Springer, Berlin, Germany, pp. 224-66.
2. Calhoun ML, Stinson AW. Integument. In: HD Dellman, EM Brown (eds.) Textbook of veterinary histology, 2nd edn. 1981, Lea and Febiger, Philadelphia, USA, pp. 378-411.
3. Tsukise A, Fujimori O, Yamada K. Histochemistry of glycoconjugates in the goat nasolabial skin with special reference to eccrine glands. Acta Anat 1988;132:150-8.[CrossRef]
4. Tsukise A, Meyer W, Fujimori O, Yamada K. The cytochemistry of glycoconjugates in the planum nasolabial glands of the goat as studied by electron microscopic methods. Histochem J 1988;20:617-23.[CrossRef][PubMed]
5. Meyer W, Bartels T. Histochemical study on the eccrine glands in the foot pad of the cat. Basic Appl Histochem 1989;33:219-38.[PubMed]
6. Meyer W, Tsukise A. Histochemistry of glycoconjugates in the skin of the bovine muzzle, with special reference to glandular structures. Acta Anat 1989;136:226-34.[CrossRef]
7. Meyer W, Tsukise A. Lectin histochemistry of snout skin and foot pads in the wolf and the domesticated dog (Mammalia: Canidae). Ann Anat 1995;177:39-49.[CrossRef]
8. Tsukise A, Meyer W, Schwarz R. Histochemistry of complex carbohydrates in the skin of the pig snout, with special reference to eccrine glands. Acta Anat 1983;115:141-50.[CrossRef]
9. Schauer R. Sialic acids: fascinating sugars in higher animals and man. Zoology 2004; 107:49-64.[CrossRef][PubMed]
10. Schauer R. Sialic acids as regulators of molecular and cellular interactions. Curr Opin Struct Biol 2009;19:507-14.[CrossRef][PubMed]
11. Stumpf P, Welsch U. Cutaneous eccrine glands of the foot pads of the Rock hyrax (Procavia capensis, Hyracoidea, Mammalia). Cells Tissues Organs 2002; 71:215-26.[CrossRef]
12. Stumpf P, Künzle H, Welsch U. Cutaneous eccrine glands of the foot pads of the small Madagascan tenrec (Echinops telfairi, Insectivora, Tenrecidae): skin glands in a primitive mammal. Cell Tissue Res 2004; 315:59-70.[CrossRef][PubMed]
13. Yasui T, Nara T, Habata I, Meyer W, Tsukise A. Functional properties of feline foot pads as studied by lectin histochemical and immunohistochemical methods. Acta Histochem 2010; 112: 169-77.[CrossRef][PubMed]
14. Schröder JM. Epithelial peptide antibiotics. Biochem Pharmacol 1999;57:121-34.[CrossRef][PubMed]
15. Bos JD, Pasch MC, Asghar SS. Defensins and complement systems from the perspective of skin immunity and autoimmunity. Clin Dermatol 2001;19:563-72.[CrossRef][PubMed]
16. Yang D, Chertov O, Oppenheim JJ. The role of mammalian antimicrobial peptides and proteins in awakening of innate host defense and adaptive immunity. Cell Mol Life Sci 2001;58:978-89.[CrossRef][PubMed]
17. Mowry RW. The special value methods that colour with acidic and vicinal hydroxyl groups in the histochemical study of mucus. With revised directions for the colloidal iron stain, the use of alcian blue 8 GX and their combination with the periodic acid-Schiff reaction. Ann NY Acad Sci 1963;106:402-23.[CrossRef]
18. Pearse AGE. Histochemistry. Theoretical and applied, vol. 2, 3rd ed. 1968, Churchill Livingstone, London, UK.
19. Ueda T, Fujimori O, Yamada K. A new histochemical method for detection of sialic acids using a physical development procedure. J Histochem Cytochem 1995;43:1045-51.[CrossRef][PubMed]
20. Spicer SS. A correlative study of the histochemical properties of rodent acid mucopolysaccharides. J Histochem Cytochem 1960;8:18-35.[CrossRef][PubMed]
21. Spicer SS, Horn RG, Leppi TJ. Histochemistry of connective tissue mucopolysaccharides. In: BM Wagner, DE Smith (eds.) The connective tissue. 1967, Williams and Wilkens, Baltimore, USA, pp. 251-303.
22. Danguy A. Perspectives in modern glycohistochemistry. Eur J Histochem 1995;39: 5-14.[PubMed]
23. Luft JH. Improvements in epoxy resin embedding methods. J Biophys Biochem Cytol 1961;9:409-14.[CrossRef][PubMed]
24. Watson ML. Staining of tissue sections for electron microscopy with heavy metals. J Biophys Biochem Cytol 1958;4:475-8.[CrossRef][PubMed]
25. Reynolds ES. The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J Cell Biol 1963;17: 208-12.[CrossRef][PubMed]
26. Newman GR, Jasani B, Williams ED. A simple post-embedding system for the rapid demonstration of tissue antigens under the electron microscope. Histochem J 1983; 15:543-55.[CrossRef][PubMed]
27. Yamada K. Histochemistry of carbohydrates as performed by physical development procedures. Histochem J 1993;25:95-106.[CrossRef][PubMed]
28. Casselman WGB. Histochemical technique. 1959, Methuen, London, UK.
29. Kurosumi K, Shibasaki S, Ito T. Cytology of the secretion in mammalian sweat glands. Int Rev Cytol 1984;87:253-329.[CrossRef]
30. Yasui T, Tsukise A, Meyer W. Histochemical analysis of glycoconjugates in the eccrine glands of the raccoon digital pads. Eur J Histochem 2004;48:393-402.[PubMed]
31. Yasui T, Tsukise A, Meyer W. Morphology and glycoconjugate histochemistry of the eccrine glands in the snout skin of the North American raccoon (Procyon lotor). Arch Dermatol Res 2005;296:482-88.[CrossRef][PubMed]
32. Morrow AL, Ruiz-Palacios GM, Jiang X, Newburg DS. Human-milk glycans that inhibit pathogen binding protect breast-feeding infants against infectious diarrhea. J Nutr 2005;135:1304-7.[PubMed]
33. Meyer W, Seegers U, Schnapper A, Neuhaus H, Himstedt W, Toepfer-Petersen E. Presence of free sugars on the epidermal surface of aquatic vertebrates and their possible involvement in antimicrobial defense. Aquatic Biol 2007;1:167-75.[PubMed]
34. Ueda T, Fujimori O, Tsukise A, Yamada K. Histochemical analysis of sialic acids in the epididymis of the rat. Histochem Cell Biol 1998;109:399-407.[PubMed]
35. Varki A, Schauer R. Sialic acids. In: A. Varki, RD Cummings, JD Esko, HH Freeze, P Stanley, CR Bertozzi, GW Hart, ME Etzler (eds.) Essentials of Glycobiology, 2nd ed. 2009, Cold Spring Harber Laboratory Press, New York, USA, pp. 199-218.
36. Illana M, Prada A, Verástegui C, González Macías MC, Fernández-Trujillo FJ. Study of the distribution of glycosidic residues in eccrine sweat glands, with special reference to the contet of sialic acid. Eur J Histochem 1997;41:41-6.[PubMed]
37. Suzuki Y. Sialobiology of influenza molecular mechanism of host range variation of influenza viruses. Biol Pharm Bull 2005; 28:399-408.[CrossRef][PubMed]
38. Parillo F, Arias MP, Supplizi AV. Glycoprofile of the different cell types present in the mucosa of the horse guttural pouches. Tissue Cell 2009;41:257-65.[CrossRef][PubMed]
39. Metze D, Jurecka W, Gebhart W, Schmidt J, Mainitz M, Niebauer G. Immunohistochemical demonstration of immunoglobulin A in human sebaceous and sweat glands. J Invest Dermatol 1989; 91: 13-7.[CrossRef]
40. Ali RS, Falconer A, Ikram M, Bissett CE, Cerio R, Quinn AG. Expression of the peptide antibiotics human β defensin-1 and human β defensin-2 in normal human skin. J Invest Dermatol 2001;117:106-11.[CrossRef][PubMed]
41. Park JH, Park GT, Cho IH, Sim SM, Yang JM, Lee DY. An antimicrobial protein, lactoferrin exists in the sweat: proteomic analysis. Exp Dermatol 2011;20:367-76.[CrossRef]
42. Ezoe K, Katsumata M. Immunohistochemical study of lysozyme in human apocrine glands. J Dermatol 1990;17:159-63.[PubMed]
43. Papini M, Simonetti S, Franceschini S, Scaringi L, Binazzi M. Lysozyme distribution in healthy human skin. Arch Dermatol Res 1982;272:167-70.[CrossRef][PubMed]
44. Duszyk M. CFTR and lysozyme secretion in human airway epithelial cells. Pflügers Arch 2001;443:S45-9.[CrossRef][PubMed]
45. Jolles P, Jolles J. What’s new in lysozyme research? Always a model system, today as yesterday. Mol Cell Biochem 1984;63:165-89.[PubMed]
46. Tang QJ, Tao KZ, Yun-Liu, Sun XJ, Geng MY, Jiang CL. Immunocytochemical localization of secretory component in Paneth cell secretory granules-rat Paneth cells participate in acquired immunity. J Mol Histol 2005;36: 331-5.[CrossRef][PubMed]
47. Snoeck V, Peters I, Cox E. The IgA system: a comparison of structure and function in different species. Vet Res 2006;37:455-67.[CrossRef][PubMed]
48. Corthésy B. Role of secretory immunoglobulin A and secretory component in the protection of mucosal surfaces. Future Microbiol 2010;5:817-29.[CrossRef][PubMed]
49. Kaetzel CS, Robinson JK, Chintalacharuvu KR, Vaerman JP, Lamm ME. The polymeric immunoglobulin receptor (secretory component) mediates transport of immune complexes across epithelial cells: a local defense function for IgA. Proc Natl Acad Sci USA 1991;88:8796-800.[CrossRef][PubMed]
50. Levay PF, Viljoen M. Lactoferrin: a general review. Haematologica 1995;80:252-67.[PubMed]
51. Valenti P, Marchetti M, Superti F, Amendolia MG, Puddu P, Gessani S, et al. Antiviral activity of lactoferrin. Adv Exp Med Biol 1998; 443:199-203.[PubMed]
52. Ward PP, Uribe-Luna S, Conneely OM. Lactoferrin and host defense. Biochem Cell Biol 2002;80:95-102.[CrossRef][PubMed]
53. Ganz T. Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol 2003; 3:710-20.[CrossRef][PubMed]
54. Ganz T. Defensins: antimicrobial peptides of vertebrates. C R Biol 2004;327:539-49.[CrossRef][PubMed]
55. Meyer W, Neurand K, Tanyolaç A. General anti-microbial properties of the integument in fleece producing sheep and goats. Small Rumin Res 2001;41:181-90.[CrossRef][PubMed]
56. Hadaway LC. Skin flora and infection. J Infus Nurs 2003; 26: 44-8.[CrossRef][PubMed]
57. Zerial M, McBride H. Rab proteins as membrane organizers. Nat Rev Mol Cell Biol 2001; 2: 107-17.[CrossRef][PubMed]
58. Millar AL, Pavlos NJ, Zheng MH. Rab3D: a regulator of exocytosis in non-neuronal cells. Histol Histopathol 2002;17:929-36.[PubMed]
59. Williams JA, Chen X, Sabbatini ME. Small G proteins as key regulators of pancreatic digestive enzyme secretion. Am J Physiol Endocrinol Metab 2009;296:E405-14.[CrossRef][PubMed]
[TOP]