Y. Zhu,1 D. Ning,1 F. Wang,1 C. Liu,1 Y. Xu,1 X. Jia,2 D. Zhu1
1Department of Endocrinology, Anhui Geriatric Institute, the First Affiliated Hospital of Anhui Medical University, Hefei; 2Comprehensive Laboratory, College of Basic Medicine, Anhui Medical University, Hefei, China
Correspondence: Dr. Defa Zhu, Department of Endocrinology, Anhui Geriatric Institute, the First Affiliated Hospital of Anhui Medical University, 218, Jixi Road, Hefei 230022, Anhui, China.
Tel. +86.551.2922338 - Fax: +86.551.5120742.
E-mail: zhudfcn@yeah.net
Dr. Xuemei Jia, Comprehensive Laboratory, College of Basic Medicine, Anhui Medical University, 69, Meishan Road, Hefei 230032, Anhui, China.
Tel. +86.551.5168340 - Fax: +86.551.5168340.
E-mail: jiaxueme@126.com
Key words: hypothyroidism, levothyroxine (T4), munc-18, syntaxin-1, hippocampus.
Acknowledgments: this study was supported by the National Natural Science Foundation of China (30772301).
Received for publication: 20 August 2011.
Accepted for publication: 12 January 2012.
©Copyright Y. Zhu et al., 2012
Licensee PAGEPress, Italy
European Journal of Histochemistry 2012; 56:e22
doi:10.4081/ejh.2012.e22
AbstractAdult-onset hypothyroidism induces a variety of impairments on hippocampus- dependent neurocognitive functioningin which many synaptic proteins in hippocampus neurons are involved. Here, we observed the effect of adult-onset hypothyroidism on the expression of syntaxin-1 and munc-18 in the dorsal hippocampus and whether the altered proteins could be restored by levothyroxine (T4) treatment. All rats were separated into 4 groups randomly: hypothyroid group, 5 μg T4 /100 g body weight (BW) treated group, 20 μg T4 /100 g BW treated group and control group. The radioimmunoassay kits were applied to assay the levels of serum T3 and T4, and the levels of syntaxin-1 and munc-18 in hippocampus were assessed by immunohistochemistry and Western blot. Both analysis corroborated that syntaxin-1 in the hypothyroid group was significantly higher. Munc-18 was lower in four layers of CA3 and dentate gyrus by immunohistochemistry. After two weeks of treatment with 5 μg T4 /100 g BW for hypothyroidism, syntaxin-1 levels were completely restored, whereas the recovery of munc-18 only located in two of the four impaired layers. Twenty μg T4 /100 g BW treatment normalized munc-18 levels. These data suggested that adult-onset hypothyroidism induced increment of syntaxin-1 and decrement of munc-18 in the dorsal hippocampus, which could be restored by T4 treatment. Larger dosage of T4 caused more effective restorations. |
Thyroid hormone insufficiency causes a variety of neurocognitive dysfunctions in adulthood brain.1,2,3,4 Previous studies revealed that adult-onset hypothyroidism induced behavioral and electrophysiological impairments of hippocampus-dependent learning and memory, in which many synaptic proteins that mediate the neuroexocytosis of hippocampus neurons were involved.5,6 Munc-18 as a key regulator of neurosecretion interacts with an N-terminal peptide sequence in syntaxin-1 to control the formation of SNARE complex.7 Syntaxin-1, together with SNAP-25 (synaptosome-associated protein of 25kDa) and VAMP-2 (vesicle- associated membrane protein-2) forms the soluble N-ethylmaleimide- sensitive factor attachment protein receptors (SNARE) complex as essential components of exocytosis.8 Docking, as the initial association of secretory vesicles with the plasma membrane, is the central element in exocytosis process and precedes formation of the SNARE complex.8 Syntaxin-1 directly promotes docking in secretory cells and docking of secretory vesicles might be syntaxin dependent.9 Munc18-1 promotes docking by promoting the formation, stability, or function of the SNARE complex.8 As it was critical for neuroexocytosis, the expressions of both proteins were not adequately studied in the brain of adult-onset hypothyroid rats.
The brain impairments induced by adult-onset hypothyroidism could be reversible through T4 replacement therapy.1,10 However, it is controversial that whether the molecular impairments in hypothyroid brain were fully restored when T4 replacement therapy returned serum T4 and T3 concentrations to the normal level.11,12,13 Previous studies showed that the hypothyroidism-induced changes of Ca2+/calmodulin- independent protein kinase (CaMKII; both phosphorylated and total levels), neurogranin, SNAP-25 and calmodulin were normalized to basal levels,5,14,15 whereas the reduced protein kinase C-γ levels in the CA1 region and synaptotagmin-1 in the CA1 or CA3 region of hippocampus were not fully restored in adult hypothyroid rats.15,16
In this study, we observed the expression levels of munc-18 and syntaxin-1, and evaluated the efficiency of different dosages of T4 replacement therapy on the altered proteins expression in dorsal hippocampus of adult-onset hypothyroidism rats.
Forty-five adult male Sprague-Dawley rats weighing 280-300 g, purchased from Nanjing Experimental Animal Center, were used in present experiments and maintained under standard laboratory conditions with a natural light-dark cycle, free access to food or water. All rats were separated into 4 groups randomly: i) H (hypothyroid) group. Eleven hypothyroid rats were obtained by daily intraperitoneal injection of 6-n-propyl-2-thiouracil (PTU), dissolved in saline solution, 1 mg/100 g body weight (BW) for six weeks; ii) T4-5 group. Twelve rats were treated with PTU for four weeks followed by daily intraperitoneal injection of low dosage of T4 (dissolved in saline solution, 5 μg/100 g BW) for two weeks;17,18 iii) T4-20 group. Twelve rats were treated according to the same protocols as the T4-5 group except increasing the dosage of T4 to 20 μg/100 g BW);19-21 iv) control (C) group. Ten control rats were treated with saline solution for six weeks. Procedures involving animals and their care were performed in accordance with the Animal Care and Use Committee of Anhui Medical University.
After the last dose delivered, all rats were anesthetized by chloral hydrate (350 mg/kg BW). The blood (1.5 mL) was collected from abdominal aorta, and immediately centrifuged at 14,000 rpm × g for 15 min.22 The serum was quickly frozen at -20°C for subsequent analysis. Serum concentrations of T3 and T4 were determined with a radioimmunoassay kit (North Institute of Biological Technology, Beijing, China). The detection ranges of the assay were 0.9-2.2 ng/mL for T3 and 45-135 ng/mL for T4.
After blood collection, the brains were dissected on ice. The right brains were fixed in 4% paraformaldehyde at 4°C for 7 days. The hippocampus from left brain was quickly isolated, evenly dissected into three parts along the longitudinal axis, and stored at -80°C for Western blot. The third part from the top of hippocampus is considered to be the dorsal hippocampus.
The fixed right hemispheres were embedded in paraffin and sectioned coronally with a microtome into 6 μm thick sections. From each rat, five sections (1 out of every 20 serial sections) of the dorsal hippocampus were selected to be mounted on polylysine-coated slides. After deparaffinization the sections were subjected to an antigen retrieval protocol by heating them in 10 mM citrate buffer (pH6.0) at 100°C for 10 min. Potential non-specific binding sites were blocked with 5% normal goat serum in PBS. The sections were then incubated with the primary polyclonal antibody, mouse anti-munc-18 (1:200; BD, Franklin Lakes, NJ, USA) or rabbit anti-syntaxin-1 (1:400; Chemicon Int., Temecula, CA, USA) at 37°C for 1 h and overnight at 4°C, followed by washes in PBS, incubation with the biotinylated secondary antibody (rabbit anti-mouse or goat anti-rabbit IgG (Maixin-Bio Ltd., Fujian, China) for 15 min at 37°C and washes in PBS. Sections were further incubated with the HRP for 10 min at 37°C, washed in PBS and colored with Diaminobenzidine (DAB; Maixin-Bio Ltd.) at room temperature for 7 min. Finally, sections were counterstained with hematoxylin for 3 min, dehydrated, rinsed, and coverslipped with glycerin. Sections that were not incubated with primary antibody served as negative controls.
An image analysis system was used for quantitative analysis.23 The system includes MetaMorph image acquisition and processing software (JADA 801D, China), a Nikon 80i microscope (Nikon, Tokio, Japan) equipped with a HP computer. The analyzed layers from different subfields of the dorsal hippocampus include the stratum oriens (SO), stratum radiatum (SR) and stratum lacunosum-moleculare (SLM) in the CA1; SO, stratum lucidum (SL) and SR in the CA3; polymorphic layer (PL) and molecular layer (ML) in the dentate gyrus (DG). First, a picture of complete hippocampal formation was obtained at low magnification of × 40. Then pictures of higher magnification of × 200 in various subfields of the hippocampus were acquired according to the size of each subfield: three pictures in CA1 for SO, SR; one picture in CA3 and DG-PL; two pictures in DG-ML and CA1-SLM. Digital data were exported into MetaMorph software for analysis and processing. The average optical density (OD) represented the intensity of immunohistochemical staining.
Crude synaptosomes from the dorsal hippocampus were prepared as previously described.24 In brief, the dorsal hippocampi were homogenized in Dounce homogenizers containing ice-cold HEPES buffer (10 mM HEPES, 1 mM EDTA, 10% sucrose, pH 7.4) and a protease inhibitor cocktail (2 μL/mL buffer; Sigma, St. Louis, MO, USA). The homogenate was centrifuged for the first time at 1000 × g for 8 min. The pellet was discarded, and the supernatant was centrifuged again at 9500 × g for 15 min. The supernatant was discarded and the pellet (crude synaptosomal fraction) was reconstituted in ice-cold HEPES buffer plus protease inhibitors as described above and stored at -80°C untill use. Protein concentration was tested by the Bio-Rad DC Protein Assay kit (Bio-Rad Laboratories). Samples (each containing 20 μg protein) were loaded on a 12% sodium dodecyl sulfate-polyacrylamide gel. Gels were run in triplicate and transferred onto a BioTrace polyvinylidene fluoride (PVDF) membrane (Amersham Biosciences, Piscataway, NJ, USA). The membrane was blocked in freshly prepared Tris-buffered saline (TBS), pH7.2, with 5% nonfat dry milk for 1 h at room temperature, then incubated with primary antibodies for munc-18 (1:1000; mouse polyclonal; BD) or syntaxin-1 (1:2500, mouse polyclonal, Sigma), overnight at 4°C, followed by wash with TBS-0.05% Tween 20 (TBS-T). The membrane was then incubated with secondary antibody [1:15,000 or 1:16,000 anti-mouse IgG-horseradish peroxidase (HRP), respectively] and HRP-conjuncted antibody for GAPDH (KangCheng, China) at room temperature for 90 min, followed by immunodetection of proteins with chemiluminescence (ECL kit; Amersham Biosciences). The protein levels of munc-18 and sytanxin-1 were determined as the relative ratio of the band intensity of protein to that of the loading control (GAPDH).
All analyses were conducted by statistical software, SPSS 17.0 for Windows. The results were expressed as mean ± means of standard error (SEM). The total serum T3, T4 levels, syntaxin-1 and munc-18 immunoreactivity of different treatment groups were determined by one-way analysis of variance (ANOVA) using least-significant difference for post hoc analysis. P<0.05 was considered significant.
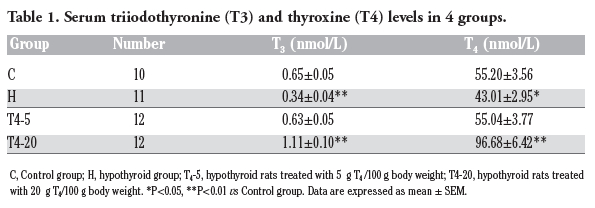
The serum T3 and T4 levels were significantly lower (P<0.05) in hypothyroid rats than those of control rats. Five μg/100 g BW T4 treatment restored T3 and T4 similar to the levels of control rats, but both of them were significantly higher (vs the C group, P<0.001) after 20 μg/100 g BW T4 treatment (Table 1).
|
Table 1. Serum triiodothyronine (T3) and thyroxine (T4) levels in 4 groups. |
Representative photomicrographs of the immuno-labeled munc-18 and syntaxin-1 in different groups were shown in Figure 1 (A, B) and Figure 2 (A, B), respectively. The distributions of the two proteins within the dorsal hippocampus were similar among the four groups. Punctate spots of reaction product distributed in every layer of CA1, CA3 and DG subfields. Large spots of munc-18 were found in the CA3-SL where large terminals of mossy fiber were located (Figures 1 and 2).
The OD values of munc-18 and syntaxin-1 immunoreactivity in each stratum of hippocampal subfields are analyzed and shown in
2 and Table 3, respectively. In the H group, the OD values of munc-18 in four layers of CA3 and DG subfields were significantly lower compared to the corresponding layers in the C group (P<0.05), i.e. CA3-SO and CA3-SR, DG-PL and DG-ML. In the T4-5 group, the OD values were significantly lower compared to the C group in DG-ML (P=0.028) and CA3-SR (P=0.091). In the T4-20 group, the OD values in all layers were similar to those in the C group.
The OD values corresponding to syntaxin-1 immunoreactivity in every layer of hippocampal subfields were significantly higher in the H group (vs the C group, P<0.001). In contrast, the OD values of T4-5 or T4-20 group were not significantly different compared to the control group in all hippocampal layers (vs the C group, P<0.05).
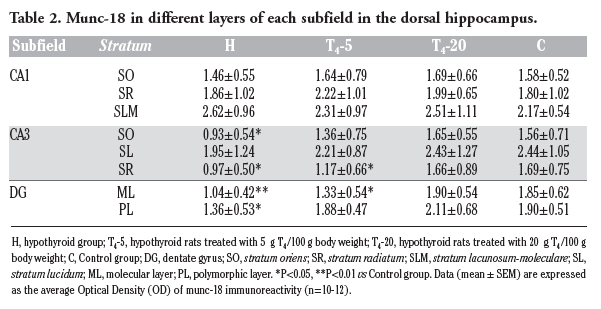 |
Table 2. Munc-18 in different layers of each subfield in the dorsal hippocampus. |
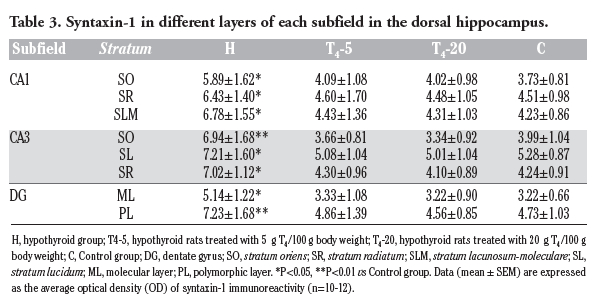 |
Table 3. Syntaxin-1 in different layers of each subfield in the dorsal hippocampus. |
The relative levels of syntaxin-1 and munc-18 in the dorsal hippocampus are shown in Figure 3. Our data showed that the amount of syntaxin-1 was increased by 36% in the hypothyroid rats (P=0.019). In the T4-5 and T4-20 groups, the amount was decreased by 25% and 38% compared to the H group and not significantly different compared to controls. As for munc-18, there were no significant differences between differently treated hypothyroid rats and control rats (P>0.05).
In the present study, a decreased expression level of munc-18 was initially corroborated in the dorsal hippocampus of rats with adult-onset hypothyroidism by immunohistochemical analyses. Thyroid hormones (TH) regulated protein synthesis in brain,25,26,27 and it has been reasoned that modifications in thyroid hormone receptors (TRs) expression by thyroid status may directly impact the expression of thyroid hormone target proteins.5,28,29 The reduced expression of munc-18 could be related to the lower TH neuronal levels and the decreased expression of TRs in hippocampus associated with hypothyroidism.30,31 Hypothyroidism may not have the same effect on syntaxin-1 expression in different regions of nervous tissue. Previous reports showed that the expression level of syntaxin-1 was downregulated in the adenohypophysis and prefrontal cortex of hypothyroid rats,14,32 however, we found that the expression of syntaxin-1 was upregulated in dorsal hippocampus of hypothyroid rats, and the mechanism underlying this is unknown. According to previous studies, hypothyroidism induced different quantitative distributions of thyroid hormones.33
These results indicated that thyroid hormones deficiency may not have the same effect on syntaxin-1 expression in different regions of nervous tissue, and the mechanism underlying this is unknown. According to previous studies, hypothyroidism induced different quantitative distributions of thyroid hormones,30 and unidentical changes of TR isoforms in different brain regions; e.g. The relative expression of TRα1 was increased and in contrast the TRα2 were decreased in hippocampus and cerebral cortex.31 It might be that syntaxin-1 was primally regulated by different TR isoforms in different nervous tissues. In this work, our results suggested that the effects of adult-onset hypothyroidism on munc-18 and syntaxin-1 are different though both proteins are required for neuroexocytosis.7,8 The binary interaction between munc18-1 and syntaxin-1 is crucial for the formation of SNARE complex required for neuroexocytosis.34 The decrement of munc-18 might not effectively buffers the levels of closed syntaxin in hippocampal neurons,35 and syntaxin-1 was overexpressed compensatory to ensure the formation of SNARE complex under insufficient munc-18.36 In the rescue experiment, the N-peptide of syntaxin-1 recruits munc-18 protein to the SNARE bundle, facilitating their assembly into a fusion-competent complex.37
T4 replacement therapy for adult-onset hypothyroidism, by which the plasma thyroid hormones reached euthyroidism, could ameliorate impaired expressions of synaptic proteins related with brain functions.1,14 In our study, syntaxin-1 levels were completely restored by T4 replacement therapy, whereas the recovery of munc-18 only located in two of the four impaired layers. By looking into previous reports, T4 replacement therapy which normalized serum T4 and T3 concentrations to the normal range fully recovered the changes of CaMKII, SNAP-25, neurogranin, and calmodulin rather than protein kinase C-γ levels in hypothyroid brain,5,14,16 suggesting that the molecular impairments in adult-onset hypothyroidism brain presented asynchronous recovery through T4 replacement therapy. It might be correlated with insufficient administrated dosage of T4. In current work, we found that the expression of munc-18 in other impaired layers was normalized and sytaxin-1 levels were more close to that of control rats through larger administrated dosage of T4. Similar effect was observed in morphological studies,38 in which the recovery of down-regulated number and distribution of dendritic spines among the apical shafts of pyramidal neurons increased with the T4 dose. As we know, thyroid hormone exerts its effects primarily through binding to thyroid hormone receptors (TRs). It might be that larger dose of T4 replacement therapy could bring a better occupancy of TRs. Recent study reported that the fractional occupancy of TRs was 49% after the multiple low doses of T3 injections and 86% after the single large dose of T3 injection, which resulted in the changed expressions of 19 genes and 237 genes respectively.39,40 Besides, the asynchronous recovery of these proteins may also suggest a protein-specific sensitivity to thyroid hormones in different neuroendocrine cells, since they had the distinct molecular properties,41,42 and the different expression patterns of thyroid hormone receptor isoforms in neurons.31
In conclusion, our results indicated that adult-onset hypothyroidism induced increment of syntaxin-1 and decrement of munc-18 in the dorsal hippocampus. T4 replacement treatment for two weeks, which normalized serum T3 and T4 levels, caused inconsistent recovery of changed syntaxin-1 and munc-18. Syntaxin-1 was fully restored while munc-18 was not. Larger dosage of T4 treatment caused more effective restorations.
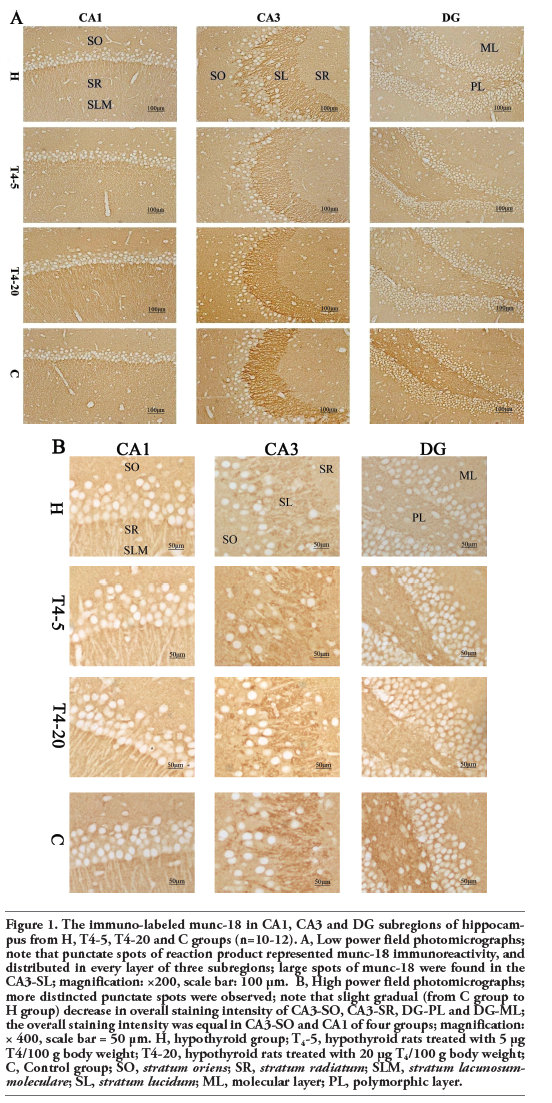 |
Figure 1. The immuno-labeled munc-18 in CA1, CA3 and DG subregions of hippocampus from H, T4-5, T4-20 and C groups (n=10-12). A, Low power field photomicrographs; note that punctate spots of reaction product represented munc-18 immunoreactivity, and distributed in every layer of three subregions; large spots of munc-18 were found in the CA3-SL; magnification: ×200, scale bar: 100 μm. B, High power field photomicrographs; more distincted punctate spots were observed; note that slight gradual (from C group to H group) decrease in overall staining intensity of CA3-SO, CA3-SR, DG-PL and DG-ML; the overall staining intensity was equal in CA3-SO and CA1 of four groups; magnification: × 400, scale bar = 50 μm. H, hypothyroid group; T4-5, hypothyroid rats treated with 5 μg T4/100 g body weight; T4-20, hypothyroid rats treated with 20 μg T4/100 g body weight; C, Control group; SO, stratum oriens; SR, stratum radiatum; SLM, stratum lacunosum-moleculare; SL, stratum lucidum; ML, molecular layer; PL, polymorphic layer. |
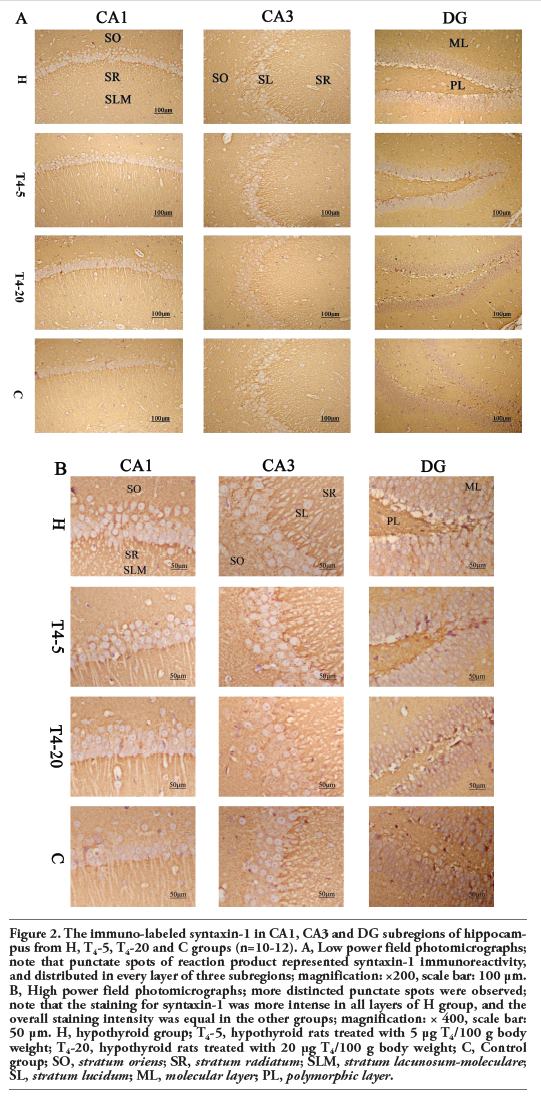 |
Figure 2. The immuno-labeled syntaxin-1 in CA1, CA3 and DG subregions of hippocampus from H, T4-5, T4-20 and C groups (n=10-12). A, Low power field photomicrographs; note that punctate spots of reaction product represented syntaxin-1 immunoreactivity, and distributed in every layer of three subregions; magnification: ×200, scale bar: 100 μm. B, High power field photomicrographs; more distincted punctate spots were observed; note that the staining for syntaxin-1 was more intense in all layers of H group, and the overall staining intensity was equal in the other groups; magnification: × 400, scale bar: 50 μm. H, hypothyroid group; T4-5, hypothyroid rats treated with 5 μg T4/100 g body weight; T4-20, hypothyroid rats treated with 20 μg T4/100 g body weight; C, Control group; SO, stratum oriens; SR, stratum radiatum; SLM, stratum lacunosum-moleculare; SL, stratum lucidum; ML, molecular layer; PL, polymorphic layer. |
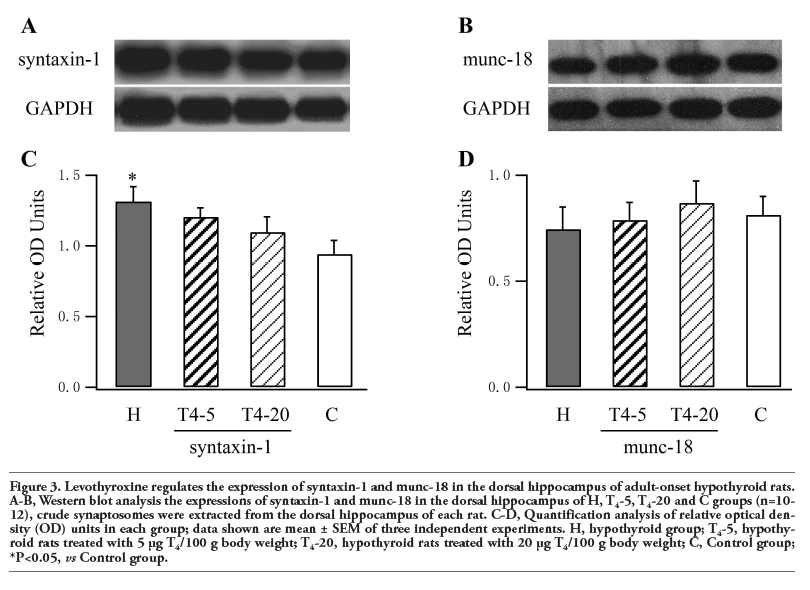 |
Figure 3. Levothyroxine regulates the expression of syntaxin-1 and munc-18 in the dorsal hippocampus of adult-onset hypothyroid rats. A-B, Western blot analysis the expressions of syntaxin-1 and munc-18 in the dorsal hippocampus of H, T4-5, T4-20 and C groups (n=10-12), crude synaptosomes were extracted from the dorsal hippocampus of each rat. C-D, Quantification analysis of relative optical density (OD) units in each group; data shown are mean ± SEM of three independent experiments. H, hypothyroid group; T4-5, hypothyroid rats treated with 5 μg T4/100 g body weight; T4-20, hypothyroid rats treated with 20 μg T4/100 g body weight; C, Control group; *P<0.05, vs Control group. |
1. Wekking EM, Appelhof BC, Fliers E, Schene AH, Huyser J, Tijssen JG, et al. Cognitive functioning and well-being in euthyroid patients on thyroxine replacement therapy for primary hypothyroidism. Eur J Endocrinol 2005;153:747-53.[CrossRef][PubMed]
2. Dugbartey AT. Neurocognitive aspects of hypothyroidism. Arch Intern Med 1998; 158:1413-8.[CrossRef][PubMed]
3. Alzoubi KH, Gerges NZ, Aleisa AM, Alkadhi KA. Levothyroxin restores hypothyroidism-induced impairment of hippocampus-dependent learning and memory: Behavioral, electrophysiological, and molecular studies. Hippocampus 2009;19: 66-78.[CrossRef][PubMed]
4. Madeira MD, Sousa N, Lima-Andrade MT, Calheiros F, Cadete-Leite A, Paula-Barbosa MM. Selective vulnerability of the hippocampal pyramidal neurons to hypothyroidism in male and female rats. J Comp Neurol 1992;322:501-18.[CrossRef][PubMed]
5. Vallortigara J, Alfos S, Micheau J, Higueret P, Enderlin V. T3 administration in adult hypothyroid mice modulates expression of proteins involved in striatal synaptic plasticity and improves motor behavior. Neurobiol Dis 2008;31:378-85.[CrossRef][PubMed]
6. Liu YF, Chen HI, Yu L, Kuo YM, Wu FS, Chuang JI, et al. Upregulation of hippocampal trkb and synaptotagmin is involved in treadmill exercise-enhanced aversive memory in mice. Neurobiol Learn Mem 2008; 90:81-9.[CrossRef][PubMed]
7. Han GA, Malintan NT, Collins BM, Meunier FA, Sugita S. Munc18-1 as a key regulator of neurosecretion. J Neurochem 2010; 115:1-10.[CrossRef][PubMed]
8. de Wit H, Walter AM, Milosevic I, Gulyás-Kovács A, Riedel D, Sørensen JB, et al. Synaptotagmin-1 docks secretory vesicles to syntaxin-1/snap-25 acceptor complexes. Cell 2009;138:935-46.[CrossRef][PubMed]
9. de Wit H, Cornelisse LN, Toonen RF, Verhage M. Docking of secretory vesicles is syntaxin dependent. PLoS One 2006;1:e126.[CrossRef][PubMed]
10. Sui L, Wang F, Li BM. Adult-onset hypothyroidism impairs paired-pulse facilitation and long-term potentiation of the rat dorsal hippocampo-medial prefrontal cortex pathway in vivo. Brain Res 2006;1096:53-60.[CrossRef][PubMed]
11. Capet C, Jego A, Denis P, Noel D, Clerc I, Cornier AC, et al. [Is cognitive change related to hypothyroidism reversible with replacement therapy?] [Article in French] Rev Med Interne 2000; 21:672-8.[CrossRef]
12. Miller KJ, Parsons TD, Whybrow PC, van Herle K, Rasgon N, van Herle A, et al. Memory improvement with treatment of hypothyroidism. Int J Neurosci 2006;116: 895-906.[CrossRef][PubMed]
13. Samuels MH, Schuff KG, Carlson NE, Carello P, Janowsky JS. Health status, psychological symptoms, mood, and cognition in l-thyroxine-treated hypothyroid subjects. Thyroid 2007;17:249-58.[CrossRef][PubMed]
14. Quintanar JL, Salinas E. Effect of hypothyroidism on synaptosomal-associated protein of 25 kda and syntaxin-1 expression in adenohypophyses of rat. J Endocrinol Invest 2002;25:754-8.[PubMed]
15. Zhu DF, Liu CL, Xu YX, Hu HL, Jia XM, Chen GH, et al. Effect of thyroxine on synaptotagmin 1 and snap-25 expression in dorsal hippocampus of adult-onset hypothyroid rats. J Endocrinol Invest 2011; 34:280-6. [PubMed]
16. Alzoubi KH, Gerges NZ, Alkadhi KA. Levothyroxin restores hypothyroidism-induced impairment of ltp of hippocampal ca1: Electrophysiological and molecular studies. Exp Neurol 2005;195:330-41.[CrossRef][PubMed]
17. Bruno AN, Diniz GP, Ricachenevsky FK, Pochmann D, Bonan CD, Barreto-Chaves ML, et al. Hypo-and hyperthyroidism affect the atp, adp and amp hydrolysis in rat hippocampal and cortical slices. Neurosci Res 2005;52:61-8.[CrossRef][PubMed]
18. Iniguez MA, Rodriguez-Pena A, Ibarrola N, Morreale de Escobar G, Bernal J. Adult rat brain is sensitive to thyroid hormone. Regulation of rc3/neurogranin mrna. J Clin Invest 1992;90:554-8.[CrossRef][PubMed]
19. Das K, Chainy GB. Modulation of rat liver mitochondrial antioxidant defence system by thyroid hormone. Biochim Biophys Acta 2001;1537:1-13.[PubMed]
20. Choudhury S, Chainy GB, Mishro MM. Experimentally induced hypo- and hyper-thyroidism influence on the antioxidant defence system in adult rat testis. Andrologia 2003;35:131-40.[CrossRef][PubMed]
21. Mutvei A, Nelson BD. The response of individual polypeptides of the mammalian respiratory chain to thyroid hormone. Arch Biochem Biophys 1989;268:215-20.[CrossRef]
22. Gerges NZ, Stringer JL, Alkadhi KA. Combination of hypothyroidism and stress abolishes early ltp in the ca1 but not dentate gyrus of hippocampus of adult rats. Brain Res 2001;922:250-60.[CrossRef]
23. Tong H, Chen GH, Liu RY, Zhou JN. Age-related learning and memory impairments in adult-onset hypothyroidism in kunming mice. Physiol Behav 2007;91:290-8.[CrossRef]
24. Magnusson KR, Scruggs B, Zhao X, Hammersmark R. Age-related declines in a two-day reference memory task are associated with changes in nmda receptor subunits in mice. BMC Neurosci 2007;8:43.[CrossRef][PubMed]
25. Sokoloff L, Klee CB. The effect of thyroid on protein synthesis in brain and other organs. Res Publ Assoc Res Nerv Ment Dis 1966;43:371-86.[PubMed]
26. Crispell KR, Parson W, Hollifield G. A study of the rate of protein synthesis before and during the administration of l-triiodothyronine to patients with myxedema and healthy volunteers using n-15 glycine. J Clin Invest 1956;35:164-9.[CrossRef][PubMed]
27. de Souza Martins SC, Romao LF, Faria JC, de Holanda Afonso RC, Murray SA, Pellizzon CH, et al. Effect of thyroid hormone t3 on myosin-va expression in the central nervous system. Brain Res 2009; 1275:1-9.[CrossRef][PubMed]
28. Shahrara S, Drvota V, Sylven C. Organ specific expression of thyroid hormone receptor mrna and protein in different human tissues. Biol Pharm Bull 1999;22:1027-33.[CrossRef][PubMed]
29. Bishop CM, McCabe CJ, Gittoes NJ, Butler PJ, Franklyn JA. Tissue-specific regulation of thyroid hormone receptor mrna isoforms and target gene proteins in domestic ducks. J Endocrinol 2000;165:607-15.[CrossRef][PubMed]
30. Broedel O, Eravci M, Fuxius S, Smolarz T, Jeitner A, Grau H, et al. Effects of hyper- and hypothyroidism on thyroid hormone concentrations in regions of the rat brain. Am J Physiol Endocrinol Metab 2003;285: E470-80.[PubMed]
31. Region-specific effects of hypothyroidism on the relative expression of thyroid hormone receptors in adult rat brain. Mol Cell Biochem 2005;278:93-100.[CrossRef][PubMed]
32. Yang HY, Sun CP, Jia XM, Gui L, Zhu DF, Ma WQ. Effect of thyroxine on snare complex and synaptotagmin-1 expression in the prefrontal cortex of rats with adult-onset hypothyroidism. J Endocrinol Invest 2012;35:312-6. [PubMed]
33. Hepp R, Grant NJ, Chasserot-Golaz S, Aunis D, Langley K. The hypophysis controls expression of snap-25 and other snares in the adrenal gland. J Neurocytol 2001;30:789-800.[CrossRef][PubMed]
34. Gulyas-Kovacs A, de Wit H, Milosevic I, Kochubey O, Toonen R, Klingauf J, et al. Munc18-1: Sequential interactions with the fusion machinery stimulate vesicle docking and priming. J Neurosci 2007;27: 8676-86.[CrossRef][PubMed]
35. Mitchell SJ, Ryan TA. Munc18-dependent regulation of synaptic vesicle exocytosis by syntaxin-1a in hippocampal neurons. Neuropharmacology 2005;48:372-80.[CrossRef][PubMed]
36. Deák F, Xu Y, Chang WP, Dulubova I, Khvotchev M, Liu X, et al. Munc18-1 binding to the neuronal snare complex controls synaptic vesicle priming. J Cell Biol 2009;184:751-64.[CrossRef][PubMed]
37. Rathore SS, Bend EG, Yu H, Hammarlund M, Jorgensen EM, Shen J. Syntaxin n-terminal peptide motif is an initiation factor for the assembly of the snare-sec1/munc18 membrane fusion complex. Proc Natl Acad Sci USA 107:22399-406.[CrossRef][PubMed]
38. Ruiz-Marcos A, Cartagena-Abella P, Martinez-Galan JR, Calvo R, Morreale de Escobar G, Escobar del Rey F. Thyroxine treatment and the recovery of pyramidal cells of the cerebral cortex from changes induced by juvenile-onset hypothyroidism. J Neurobiol 1994;25:808-18.[CrossRef][PubMed]
39. Oppenheimer JH, Coulombe P, Schwartz HL, Gutfeld NW. Nonlinear (amplified) relationship between nuclear occupancy by triiodothyronine and the appearance rate of hepatic alpha-glycerophosphate dehydrogenase and malic enzyme in the rat. J Clin Invest 1978;61:987-97.[CrossRef][PubMed]
40. Diez D, Grijota-Martinez C, Agretti P, De Marco G, Tonacchera M, Pinchera A, et al. Thyroid hormone action in the adult brain: Gene expression profiling of the effects of single and multiple doses of triiodo-l-thyronine in the rat striatum. Endocrinology 2008;149:3989-4000.[CrossRef][PubMed]
41. Brunger AT. Structure of proteins involved in synaptic vesicle fusion in neurons. Annu Rev Biophys Biomol Struct 2001;30: 157-71.[CrossRef][PubMed]
42. Hay JC. Snare complex structure and function. Exp Cell Res 2001;271:10-21.[CrossRef][PubMed]
[TOP]